

Venom Peptides with Dual Modulatory Activity on the Voltage-Gated Sodium Channel NaV1.1 Provide Novel Leads for Development of Antiepileptic Drugs
- PMID: 32259093
- PMCID: PMC7088997
- DOI: 10.1021/acsptsci.9b00079
Venom Peptides with Dual Modulatory Activity on the Voltage-Gated Sodium Channel NaV1.1 Provide Novel Leads for Development of Antiepileptic Drugs
Abstract
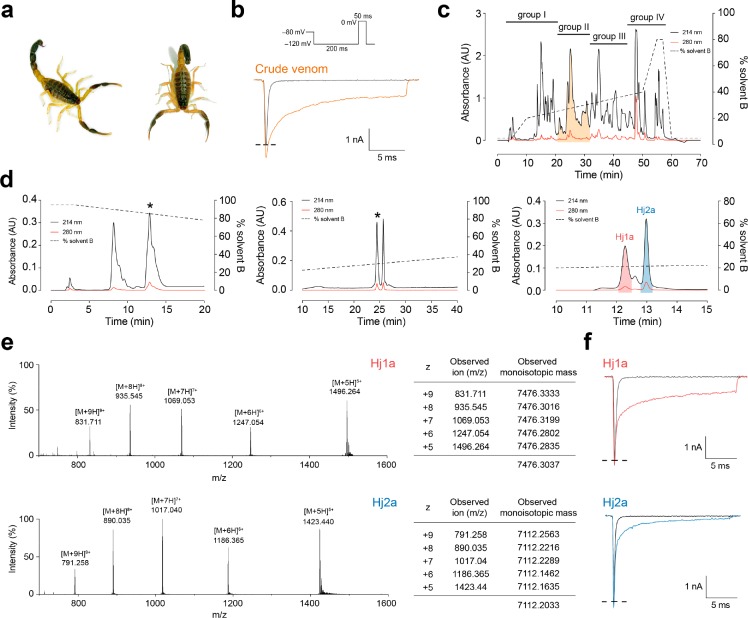
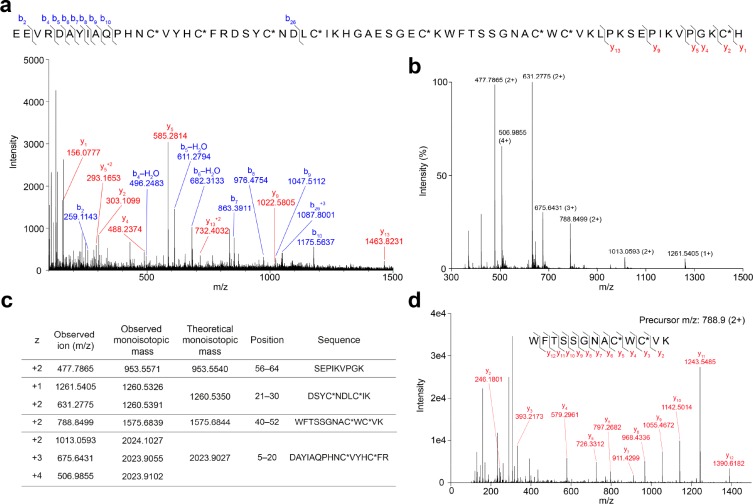
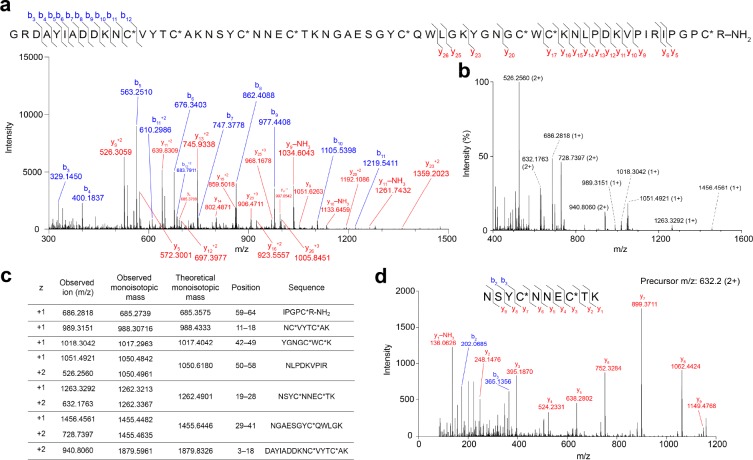
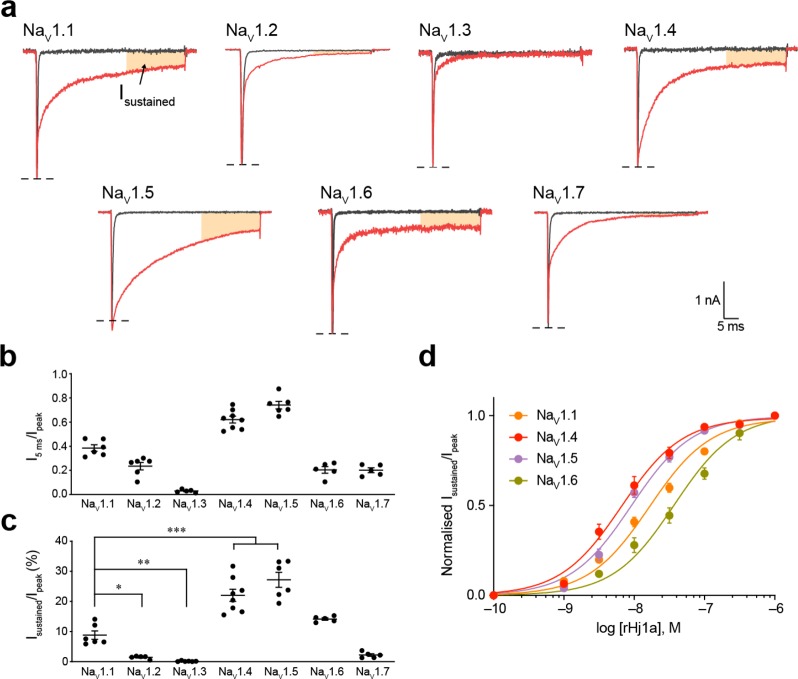
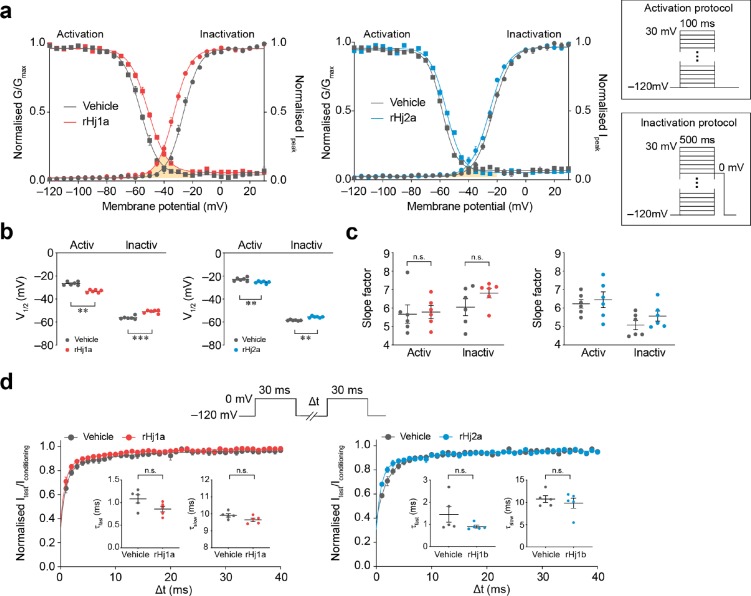
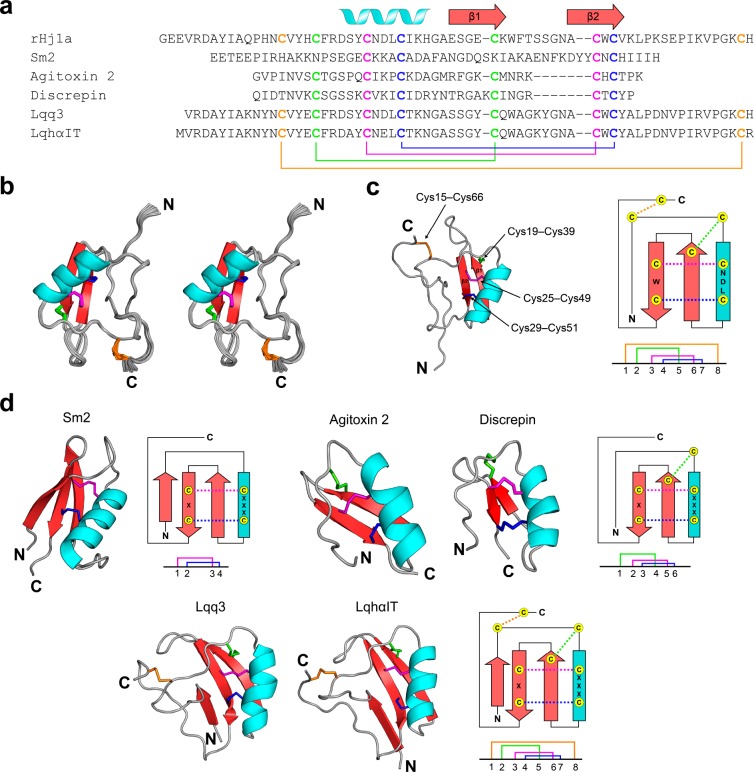
Voltage-gated sodium (NaV) channels play a fundamental role in normal neurological function, especially via the initiation and propagation of action potentials. The NaV1.1 subtype is found in inhibitory interneurons of the brain and it is essential for maintaining a balance between excitation and inhibition in neuronal networks. Heterozygous loss-of-function mutations of SCN1A, the gene encoding NaV1.1, underlie Dravet syndrome (DS), a severe pediatric epilepsy. We recently demonstrated that selective inhibition of NaV1.1 inactivation prevents seizures and premature death in a mouse model of DS. Thus, selective modulators of NaV1.1 might be useful therapeutics for treatment of DS as they target the underlying molecular deficit. Numerous scorpion-venom peptides have been shown to modulate the activity of NaV channels, but little is known about their activity at NaV1.1. Here we report the isolation, sequence, three-dimensional structure, recombinant production, and functional characterization of two peptidic modulators of NaV1.1 from venom of the buthid scorpion Hottentotta jayakari. These peptides, Hj1a and Hj2a, are potent agonists of NaV1.1 (EC50 of 17 and 32 nM, respectively), and they present dual α/β activity by modifying both the activation and inactivation properties of the channel. NMR studies of rHj1a indicate that it adopts a cystine-stabilized αβ fold similar to known scorpion toxins. Although Hj1a and Hj2a have only limited selectivity for NaV1.1, their unusual dual mode of action provides an alternative approach to the development of selective NaV1.1 modulators for the treatment of DS.
Copyright © 2019 American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures


References
-
- Verret L.; Mann E. O.; Hang G. B.; Barth A. M.; Cobos I.; Ho K.; Devidze N.; Masliah E.; Kreitzer A. C.; Mody I.; Mucke L.; Palop J. J. (2012) Inhibitory interneuron deficit links altered network activity and cognitive dysfunction in Alzheimer model. Cell 149, 708–721. 10.1016/j.cell.2012.02.046. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources