

Extraordinary Sequence Diversity and Promiscuity of Centromeric Satellites in the Legume Tribe Fabeae
- PMID: 32259249
- PMCID: PMC7403623
- DOI: 10.1093/molbev/msaa090
Extraordinary Sequence Diversity and Promiscuity of Centromeric Satellites in the Legume Tribe Fabeae
Abstract
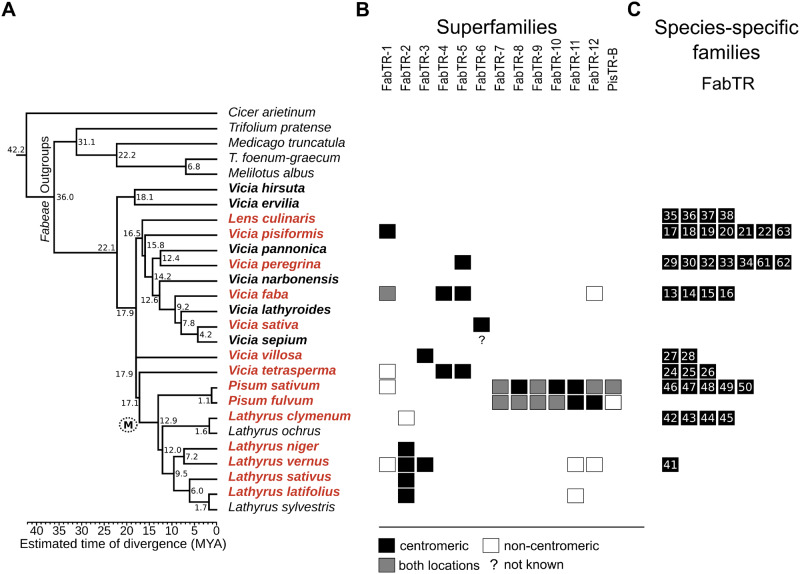
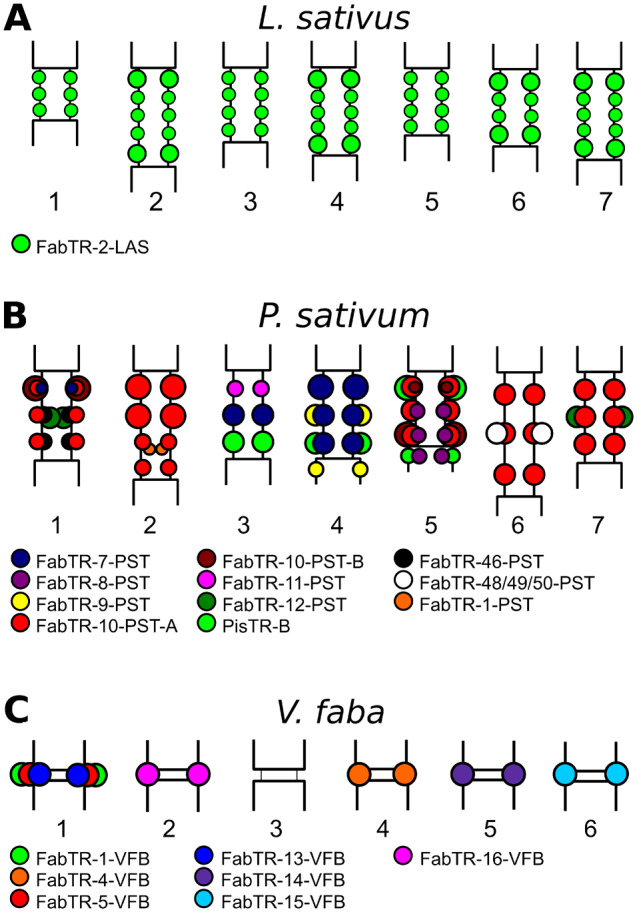
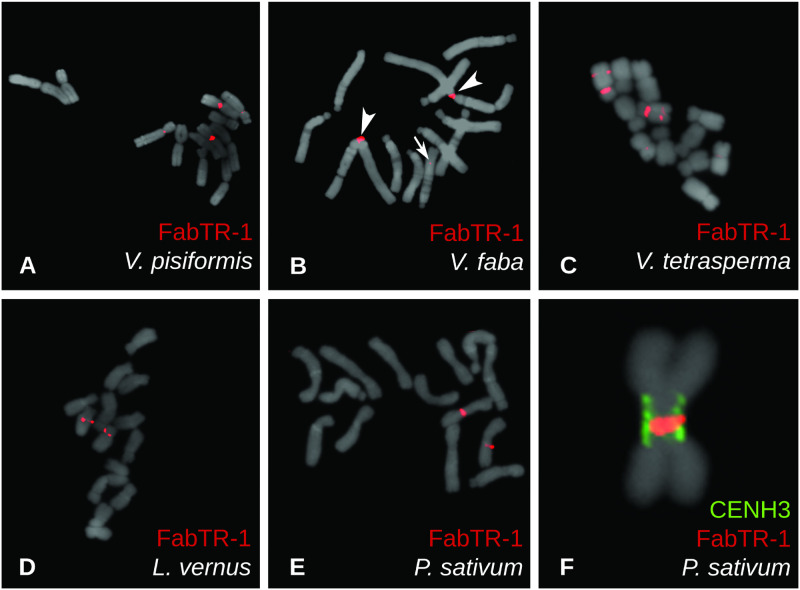
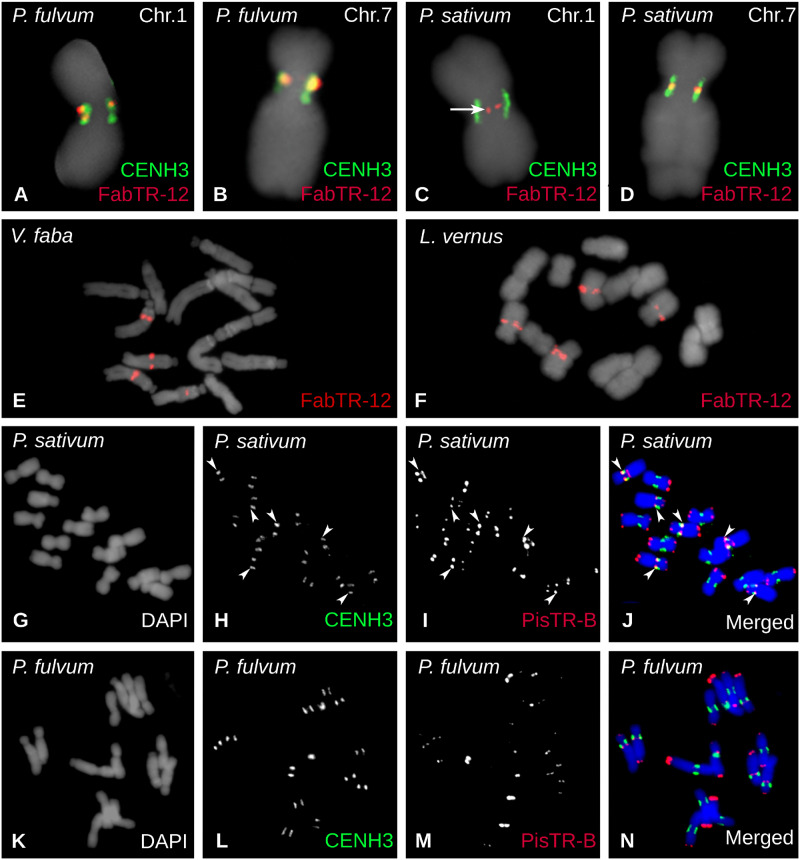
Satellite repeats are major sequence constituents of centromeres in many plant and animal species. Within a species, a single family of satellite sequences typically occupies centromeres of all chromosomes and is absent from other parts of the genome. Due to their common origin, sequence similarities exist among the centromere-specific satellites in related species. Here, we report a remarkably different pattern of centromere evolution in the plant tribe Fabeae, which includes genera Pisum, Lathyrus, Vicia, and Lens. By immunoprecipitation of centromeric chromatin with CENH3 antibodies, we identified and characterized a large and diverse set of 64 families of centromeric satellites in 14 species. These families differed in their nucleotide sequence, monomer length (33-2,979 bp), and abundance in individual species. Most families were species-specific, and most species possessed multiple (2-12) satellites in their centromeres. Some of the repeats that were shared by several species exhibited promiscuous patterns of centromere association, being located within CENH3 chromatin in some species, but apart from the centromeres in others. Moreover, FISH experiments revealed that the same family could assume centromeric and noncentromeric positions even within a single species. Taken together, these findings suggest that Fabeae centromeres are not shaped by the coevolution of a single centromeric satellite with its interacting CENH3 proteins, as proposed by the centromere drive model. This conclusion is also supported by the absence of pervasive adaptive evolution of CENH3 sequences retrieved from Fabeae species.
Keywords: CENH3; ChIP-seq; centromere evolution; plant chromosomes; satellite DNA.
© The Author(s) 2020. Published by Oxford University Press on behalf of the Society for Molecular Biology and Evolution.
Figures
References
-
- Badr SF. 2006. Karyotype analysis and chromosome evolution in species of Lathyrus (Fabaceae). Cytologia 71(4):447–455. - PubMed
-
- Bilinski P, Distor K, Gutierrez-Lopez J, Mendoza Mendoza G, Shi J, Dawe RK, Ross-Ibarra J.. 2015. Diversity and evolution of centromere repeats in the maize genome. Chromosoma 124(1):57–65. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
