

Postingestive Modulation of Food Seeking Depends on Vagus-Mediated Dopamine Neuron Activity
- PMID: 32259476
- PMCID: PMC7710496
- DOI: 10.1016/j.neuron.2020.03.009
Postingestive Modulation of Food Seeking Depends on Vagus-Mediated Dopamine Neuron Activity
Erratum in
-
Postingestive Modulation of Food Seeking Depends on Vagus-Mediated Dopamine Neuron Activity.Neuron. 2023 Feb 15;111(4):595. doi: 10.1016/j.neuron.2023.01.025. Neuron. 2023. PMID: 36796329 Free PMC article. No abstract available.
Abstract
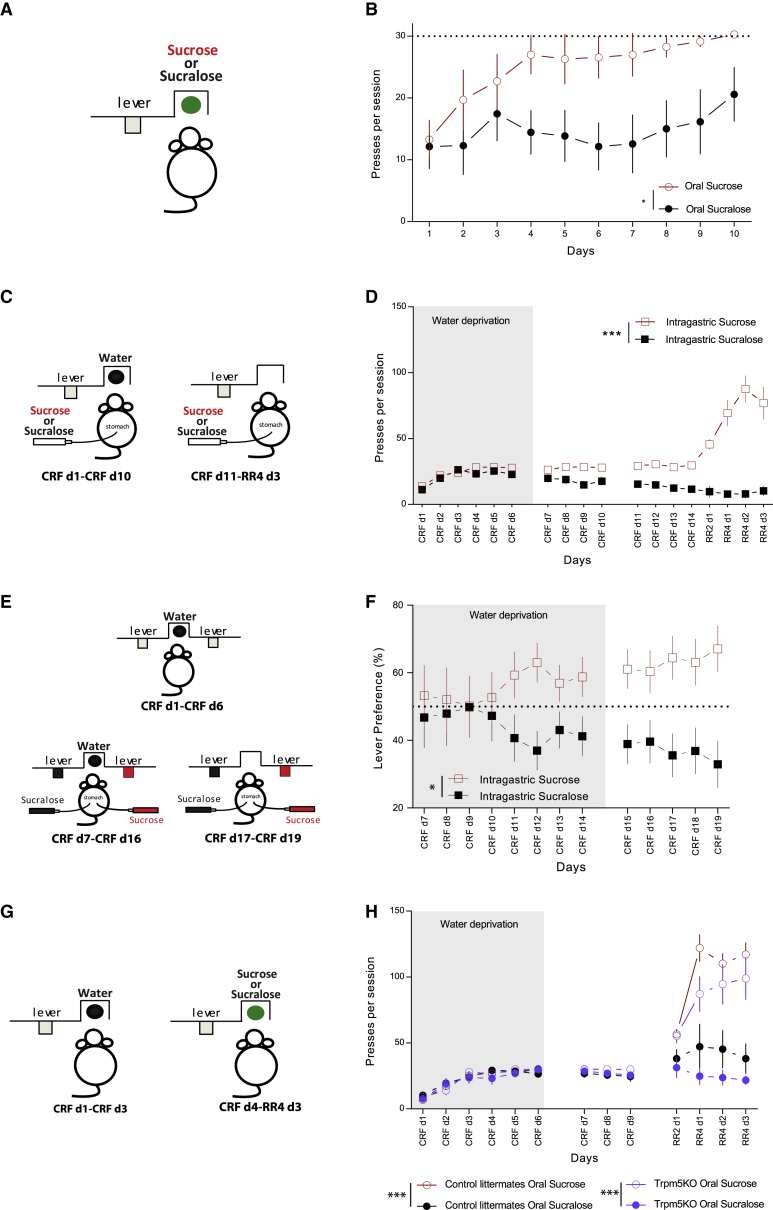
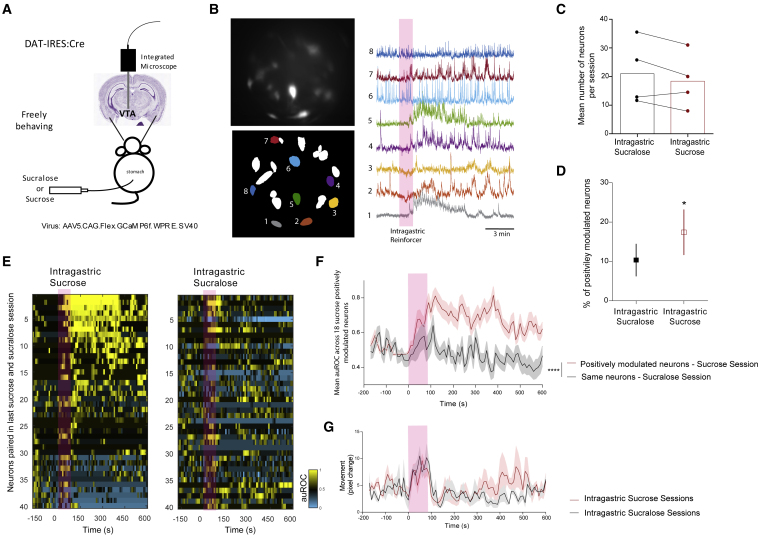
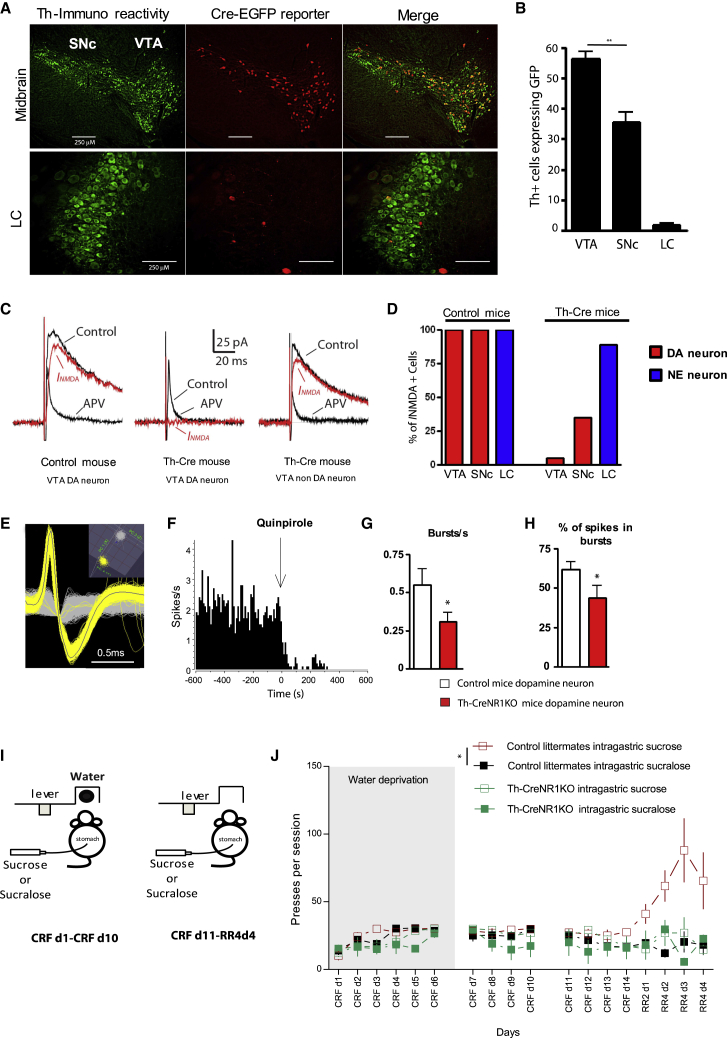
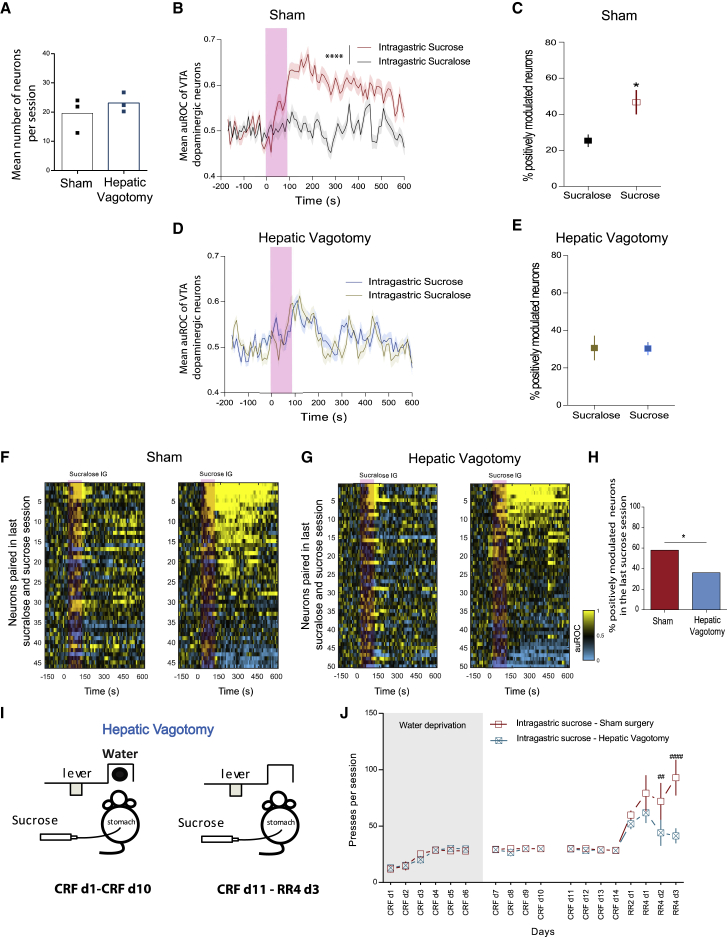
Postingestive nutrient sensing can induce food preferences. However, much less is known about the ability of postingestive signals to modulate food-seeking behaviors. Here we report a causal connection between postingestive sucrose sensing and vagus-mediated dopamine neuron activity in the ventral tegmental area (VTA), supporting food seeking. The activity of VTA dopamine neurons increases significantly after administration of intragastric sucrose, and deletion of the NMDA receptor in these neurons, which affects bursting and plasticity, abolishes lever pressing for postingestive sucrose delivery. Furthermore, lesions of the hepatic branch of the vagus nerve significantly impair postingestive-dependent VTA dopamine neuron activity and food seeking, whereas optogenetic stimulation of left vagus nerve neurons significantly increases VTA dopamine neuron activity. These data establish a necessary role of vagus-mediated dopamine neuron activity in postingestive-dependent food seeking, which is independent of taste signaling.
Keywords: Postingestive; Ventral Tegmental Area; dopamine; feeding behavior; food seeking; nutrient sensing; vagus nerve.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests A.J.O.-M. is the recipient of a grant from Schuhfried GmBH for norming and validation of cognitive tests and the national coordinator for Portugal of a non-interventional Study (EDMS-ERI-143085581, 4.0) sponsored by Janssen-Cilag Ltd., both outside of this work.
Figures
Comment in
-
A Gut Feeling about Dopamine.Neuron. 2020 Jun 3;106(5):703-704. doi: 10.1016/j.neuron.2020.05.018. Neuron. 2020. PMID: 32497503
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
