

Comparative profiling and comprehensive quantification of stratum corneum ceramides in humans and mice by LC/MS/MS
- PMID: 32265320
- PMCID: PMC7269764
- DOI: 10.1194/jlr.RA120000671
Comparative profiling and comprehensive quantification of stratum corneum ceramides in humans and mice by LC/MS/MS
Abstract
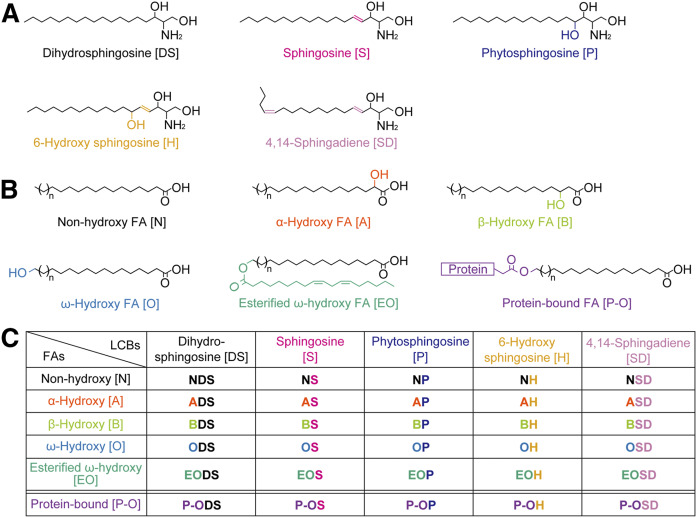
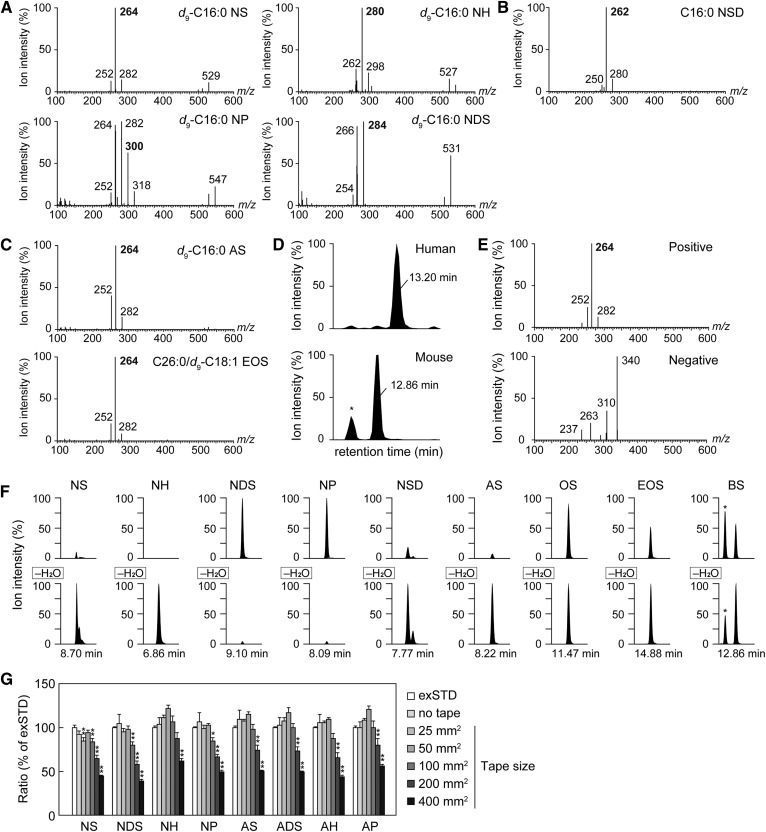
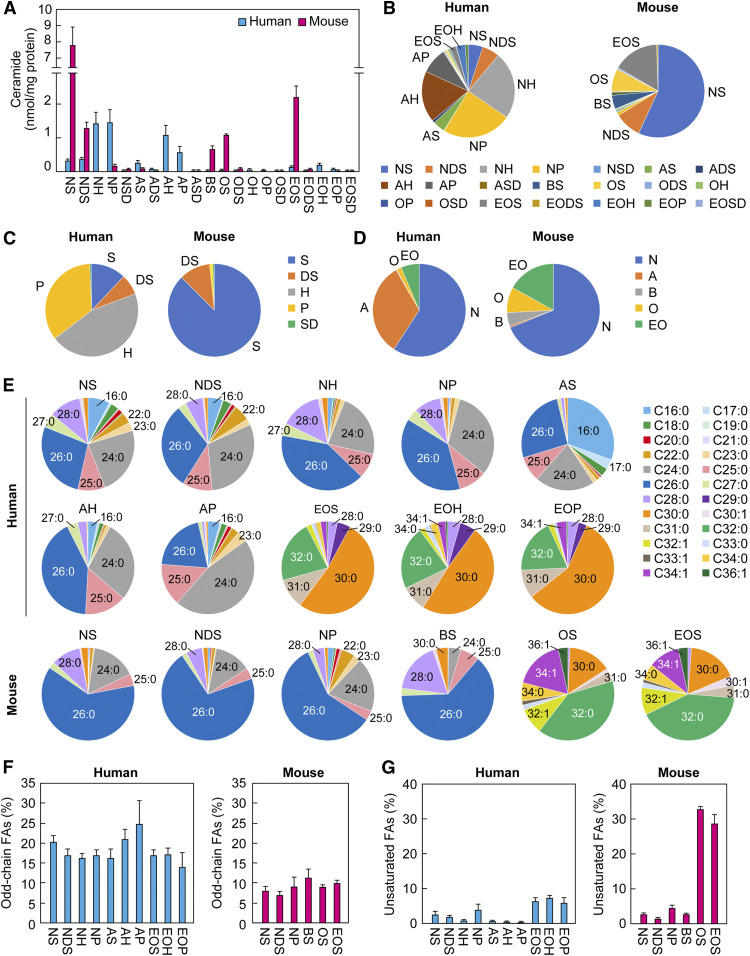
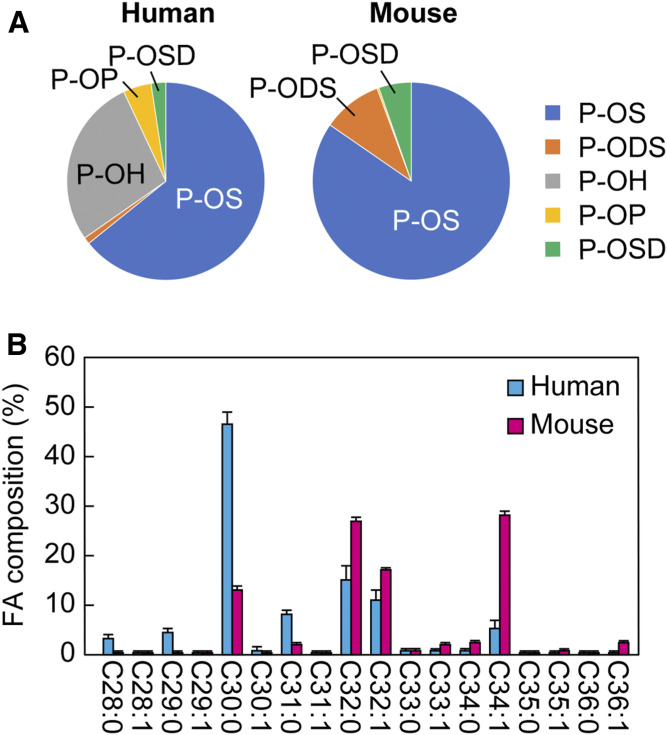
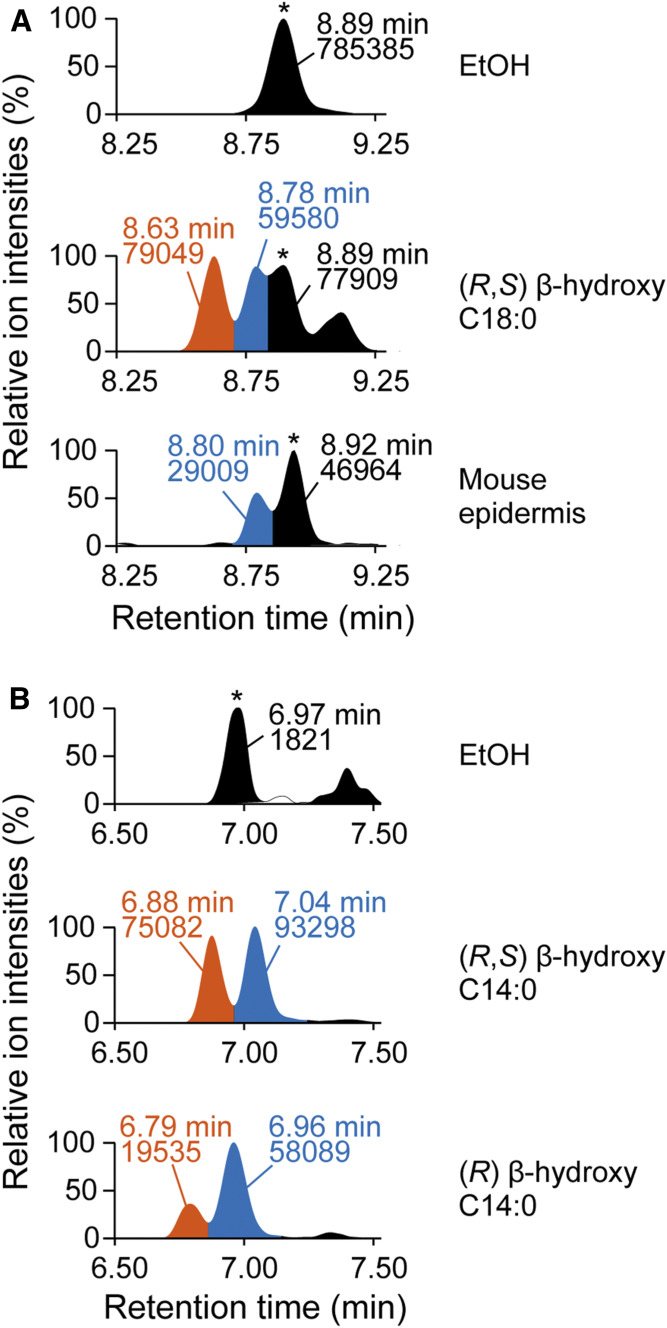
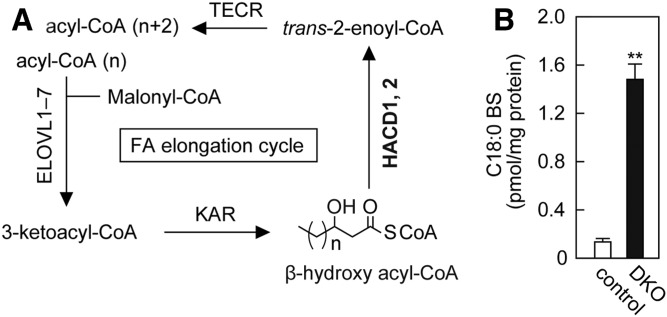
Ceramides are the predominant lipids in the stratum corneum (SC) and are crucial components for normal skin barrier function. Although the composition of various ceramide classes in the human SC has been reported, that in mice is still unknown, despite mice being widely used as animal models of skin barrier function. Here, we performed LC/MS/MS analyses using recently available ceramide class standards to measure 25 classes of free ceramides and 5 classes of protein-bound ceramides from human and mouse SC. Phytosphingosine- and 6-hydroxy sphingosine-type ceramides, which both contain an additional hydroxyl group, were abundant in the human SC (35% and 45% of total ceramides, respectively). In contrast, in mice, phytosph-ingosine- and 6-hydroxy sphingosine-type ceramides were present at ∼1% and undetectable levels, respectively, and sphingosine-type ceramides accounted for ∼90%. In humans, ceramides containing α-hydroxy FA were abundant, whereas ceramides containing β-hydroxy or ω-hydroxy FA were abundant in mice. The hydroxylated β-carbon in β-hydroxy ceramides was in the (R) configuration. Genetic knockout of β-hydroxy acyl-CoA dehydratases in HAP1 cells increased β-hydroxy ceramide levels, suggesting that β-hydroxy acyl-CoA, an FA-elongation cycle intermediate in the ER, is a substrate for β-hydroxy ceramide synthesis. We anticipate that our methods and findings will help to elucidate the role of each ceramide class in skin barrier formation and in the pathogenesis of skin disorders.
Keywords: epidermis; fatty acid; lipidomics; mass spectrometry; skin barrier; sphingolipids.
Copyright © 2020 Kawana et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
-
- Traupe H., Fischer J., and Oji V.. 2014. Nonsyndromic types of ichthyoses – an update. J. Dtsch. Dermatol. Ges. 12: 109–121. - PubMed
-
- Yardley H. J., and Summerly R.. 1981. Lipid composition and metabolism in normal and diseased epidermis. Pharmacol. Ther. 13: 357–383. - PubMed
-
- Masukawa Y., Narita H., Shimizu E., Kondo N., Sugai Y., Oba T., Homma R., Ishikawa J., Takagi Y., Kitahara T., et al. 2008. Characterization of overall ceramide species in human stratum corneum. J. Lipid Res. 49: 1466–1476. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
