

ZBTB1 Regulates Asparagine Synthesis and Leukemia Cell Response to L-Asparaginase
- PMID: 32268116
- PMCID: PMC7219601
- DOI: 10.1016/j.cmet.2020.03.008
ZBTB1 Regulates Asparagine Synthesis and Leukemia Cell Response to L-Asparaginase
Abstract
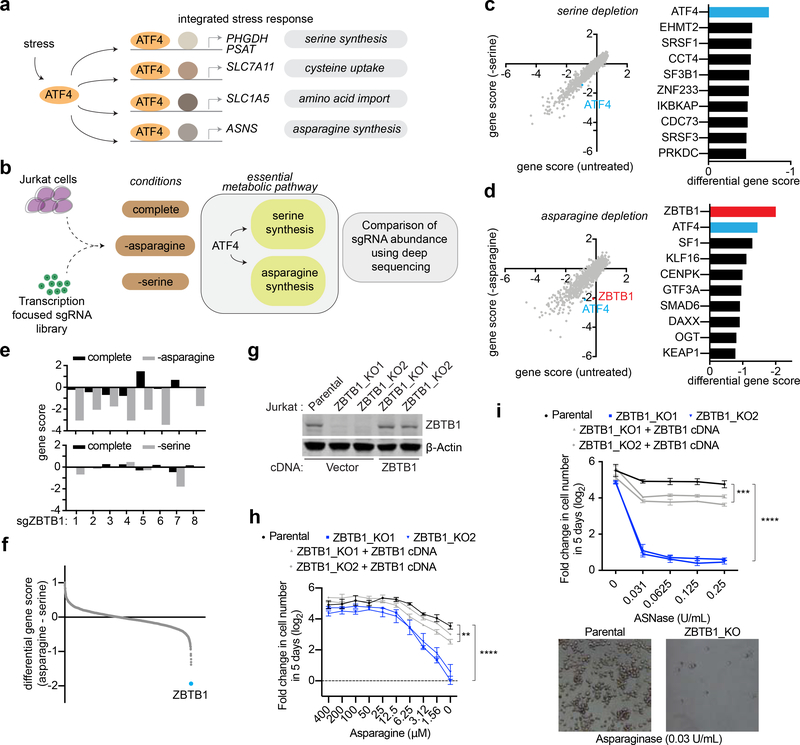
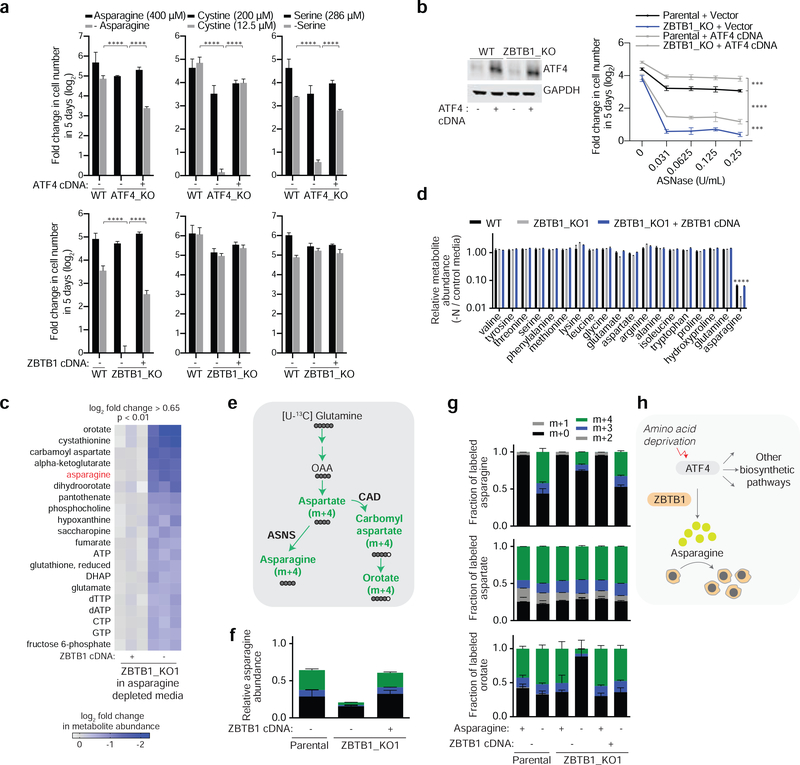
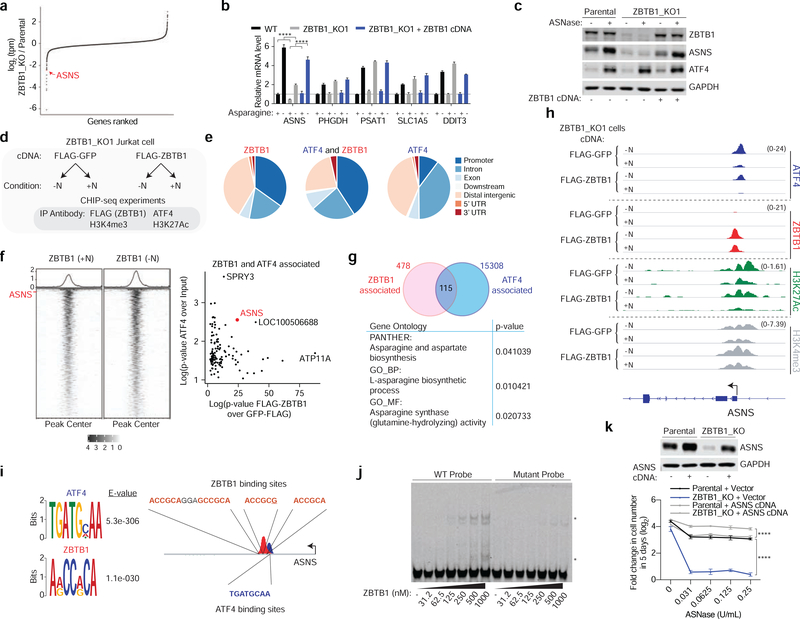
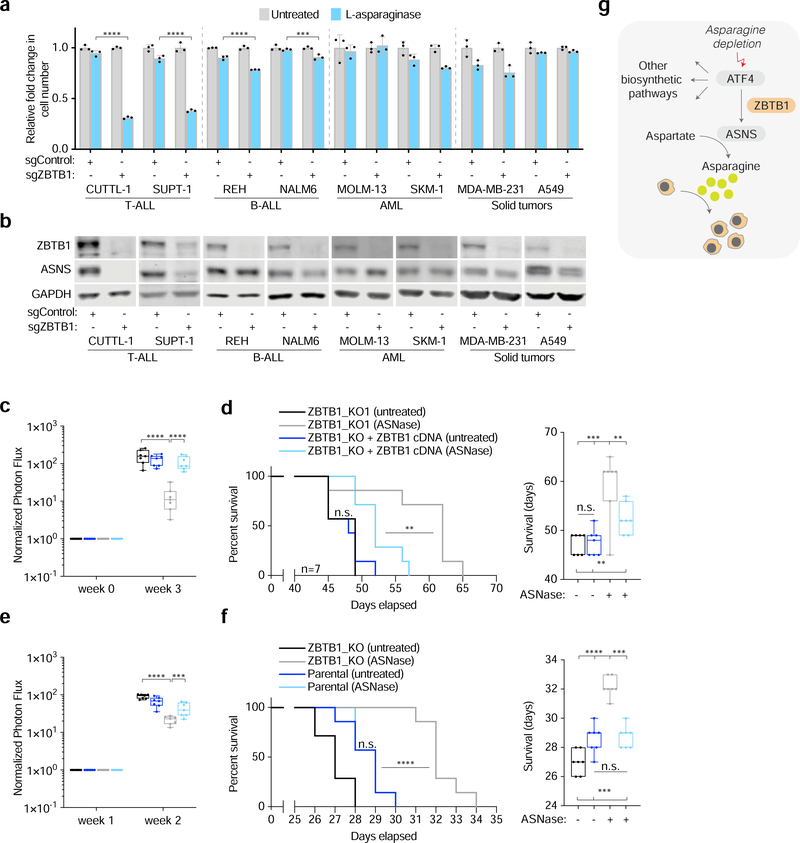
Activating transcription factor 4 (ATF4) is a master transcriptional regulator of the integrated stress response (ISR) that enables cell survival under nutrient stress. The mechanisms by which ATF4 couples metabolic stresses to specific transcriptional outputs remain unknown. Using functional genomics, we identified transcription factors that regulate the responses to distinct amino acid deprivation conditions. While ATF4 is universally required under amino acid starvation, our screens yielded a transcription factor, Zinc Finger and BTB domain-containing protein 1 (ZBTB1), as uniquely essential under asparagine deprivation. ZBTB1 knockout cells are unable to synthesize asparagine due to reduced expression of asparagine synthetase (ASNS), the enzyme responsible for asparagine synthesis. Mechanistically, ZBTB1 binds to the ASNS promoter and promotes ASNS transcription. Finally, loss of ZBTB1 sensitizes therapy-resistant T cell leukemia cells to L-asparaginase, a chemotherapeutic that depletes serum asparagine. Our work reveals a critical regulator of the nutrient stress response that may be of therapeutic value.
Keywords: ATF4; CRISPR; asparaginase; cancer metabolism; genetic screen; leukemia; transcription.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
