

Advanced Resistance Studies Identify Two Discrete Mechanisms in Staphylococcus aureus to Overcome Antibacterial Compounds that Target Biotin Protein Ligase
- PMID: 32268615
- PMCID: PMC7235819
- DOI: 10.3390/antibiotics9040165
Advanced Resistance Studies Identify Two Discrete Mechanisms in Staphylococcus aureus to Overcome Antibacterial Compounds that Target Biotin Protein Ligase
Abstract
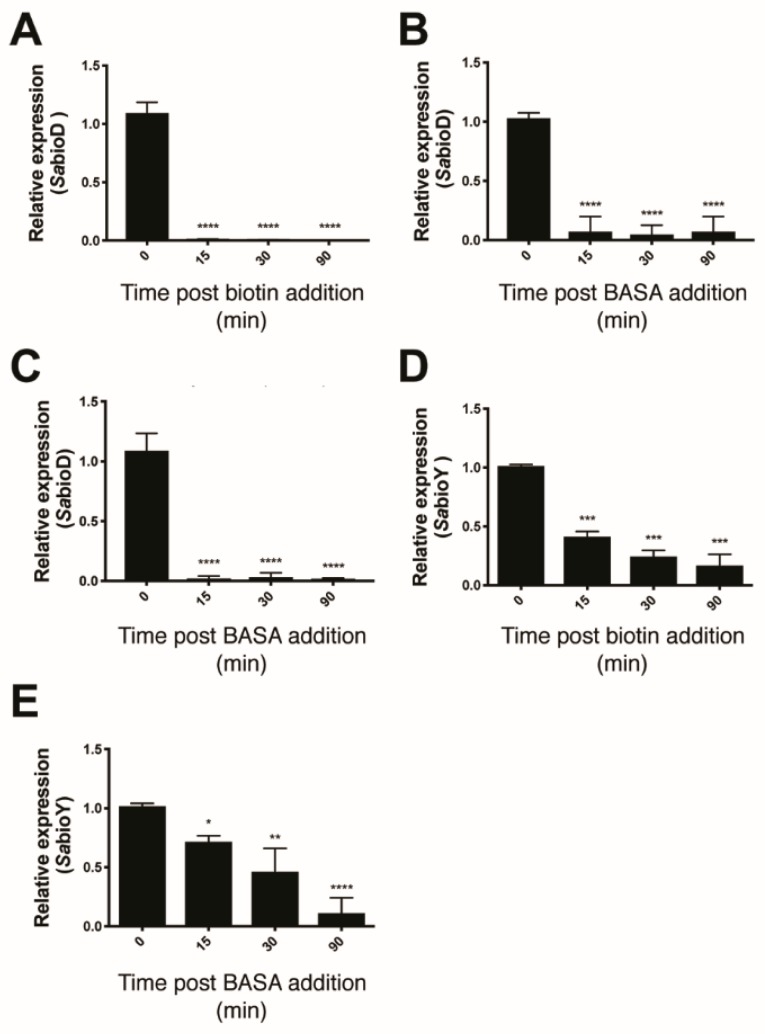
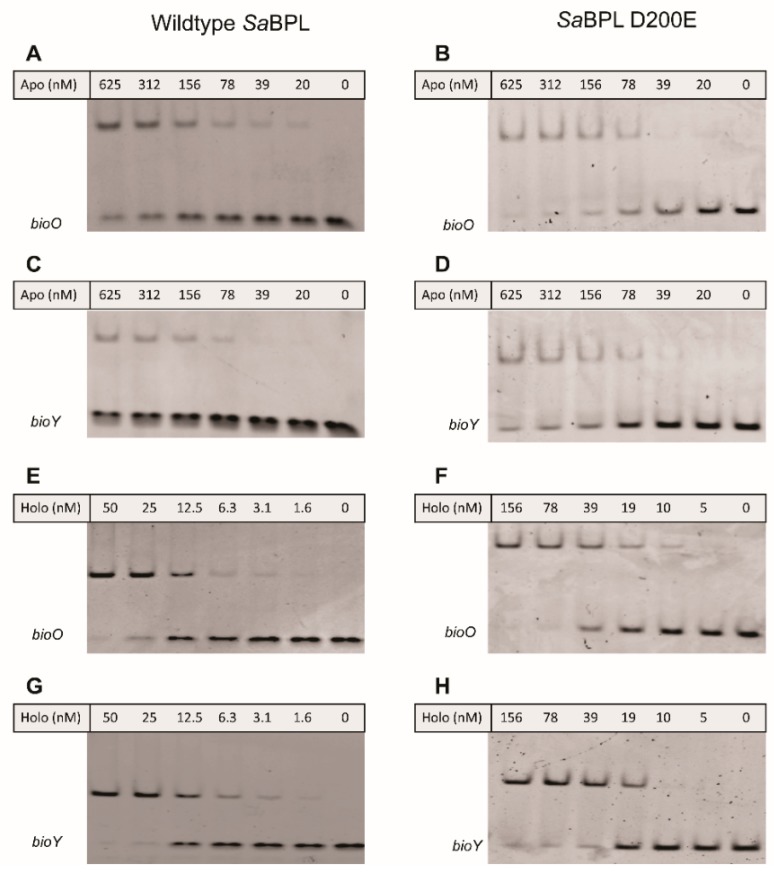
Biotin protein ligase (BPL) inhibitors are a novel class of antibacterial that target clinically important methicillin-resistant Staphylococcus aureus (S. aureus). In S. aureus, BPL is a bifunctional protein responsible for enzymatic biotinylation of two biotin-dependent enzymes, as well as serving as a transcriptional repressor that controls biotin synthesis and import. In this report, we investigate the mechanisms of action and resistance for a potent anti-BPL, an antibacterial compound, biotinyl-acylsulfamide adenosine (BASA). We show that BASA acts by both inhibiting the enzymatic activity of BPL in vitro, as well as functioning as a transcription co-repressor. A low spontaneous resistance rate was measured for the compound (<10-9) and whole-genome sequencing of strains evolved during serial passaging in the presence of BASA identified two discrete resistance mechanisms. In the first, deletion of the biotin-dependent enzyme pyruvate carboxylase is proposed to prioritize the utilization of bioavailable biotin for the essential enzyme acetyl-CoA carboxylase. In the second, a D200E missense mutation in BPL reduced DNA binding in vitro and transcriptional repression in vivo. We propose that this second resistance mechanism promotes bioavailability of biotin by derepressing its synthesis and import, such that free biotin may outcompete the inhibitor for binding BPL. This study provides new insights into the molecular mechanisms governing antibacterial activity and resistance of BPL inhibitors in S. aureus.
Keywords: BirA; Gram-positive bacteria; Staphylococcus aureus; advanced resistance studies; antimicrobial resistance; biotin; biotin protein ligase; novel antibacterials.
Conflict of interest statement
The authors declare no competing financial interest.
Figures



References
-
- Rushton J., Stärk K.D.C., Ferreira J.P. Antimicrobial Resistance. Organisation for Economic Co-Operation and Development (OECD); Paris, France: 2014.
-
- World Health Organisation . Global Action Plan on Antimicrobial Resistance. WHO; Geneva, Switzerland: 2015.
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
