

Swine ANP32A Supports Avian Influenza Virus Polymerase
- PMID: 32269123
- PMCID: PMC7307101
- DOI: 10.1128/JVI.00132-20
Swine ANP32A Supports Avian Influenza Virus Polymerase
Abstract
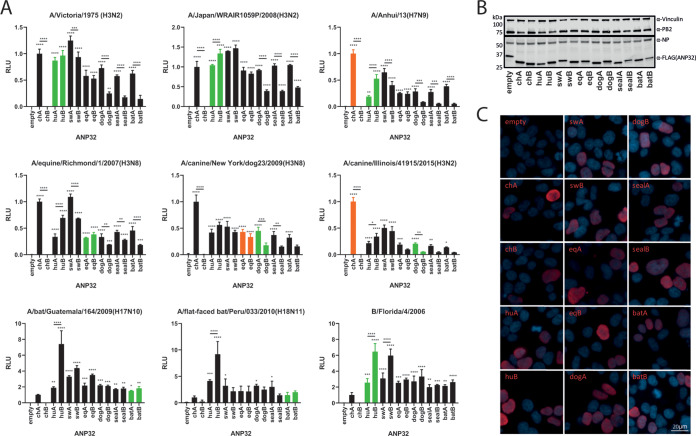
Avian influenza viruses occasionally infect and adapt to mammals, including humans. Swine are often described as "mixing vessels," being susceptible to both avian- and human-origin viruses, which allows the emergence of novel reassortants, such as the precursor to the 2009 H1N1 pandemic. ANP32 proteins are host factors that act as influenza virus polymerase cofactors. In this study, we describe how swine ANP32A, uniquely among the mammalian ANP32 proteins tested, supports the activity of avian-origin influenza virus polymerases and avian influenza virus replication. We further show that after the swine-origin influenza virus emerged in humans and caused the 2009 pandemic, it evolved polymerase gene mutations that enabled it to more efficiently use human ANP32 proteins. We map the enhanced proviral activity of swine ANP32A to a pair of amino acids, 106 and 156, in the leucine-rich repeat and central domains and show these mutations enhance binding to influenza virus trimeric polymerase. These findings help elucidate the molecular basis for the mixing vessel trait of swine and further our understanding of the evolution and ecology of viruses in this host.IMPORTANCE Avian influenza viruses can jump from wild birds and poultry into mammalian species such as humans or swine, but they only continue to transmit if they accumulate mammalian adapting mutations. Pigs appear uniquely susceptible to both avian and human strains of influenza and are often described as virus "mixing vessels." In this study, we describe how a host factor responsible for regulating virus replication, ANP32A, is different between swine and humans. Swine ANP32A allows a greater range of influenza viruses, specifically those from birds, to replicate. It does this by binding the virus polymerase more tightly than the human version of the protein. This work helps to explain the unique properties of swine as mixing vessels.
Keywords: ANP32; ANP32A; ANP32B; host factors; influenza; pandemic; replication; swine; swine influenza; zoonotic.
Copyright © 2020 Peacock et al.
Figures
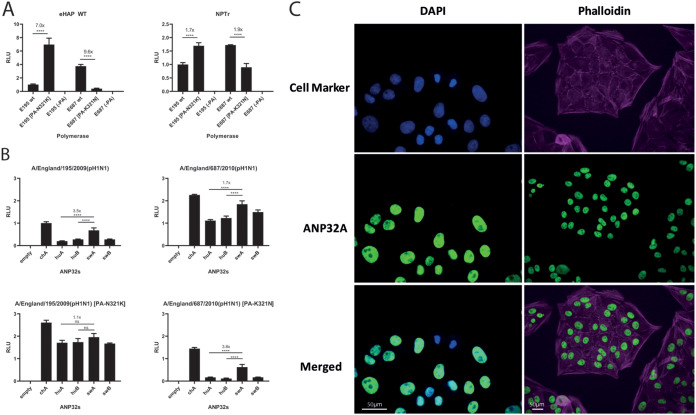

References
-
- Kandeil A, Gomaa MR, Shehata MM, El Taweel AN, Mahmoud SH, Bagato O, Moatasim Y, Kutkat O, Kayed AS, Dawson P, Qiu X, Bahl J, Webby RJ, Karesh WB, Kayali G, Ali MA. 2018. Isolation and Characterization of a Distinct Influenza A Virus from Egyptian Bats. J Virol 93:e01059-18. doi:10.1128/JVI.01059-18. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 205100/WT_/Wellcome Trust/United Kingdom
- BB/S008292/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/P013740/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/K002465/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/R013071/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
