

DNA clamp function of the monoubiquitinated Fanconi anaemia ID complex
- PMID: 32269332
- PMCID: PMC7398534
- DOI: 10.1038/s41586-020-2110-6
DNA clamp function of the monoubiquitinated Fanconi anaemia ID complex
Abstract
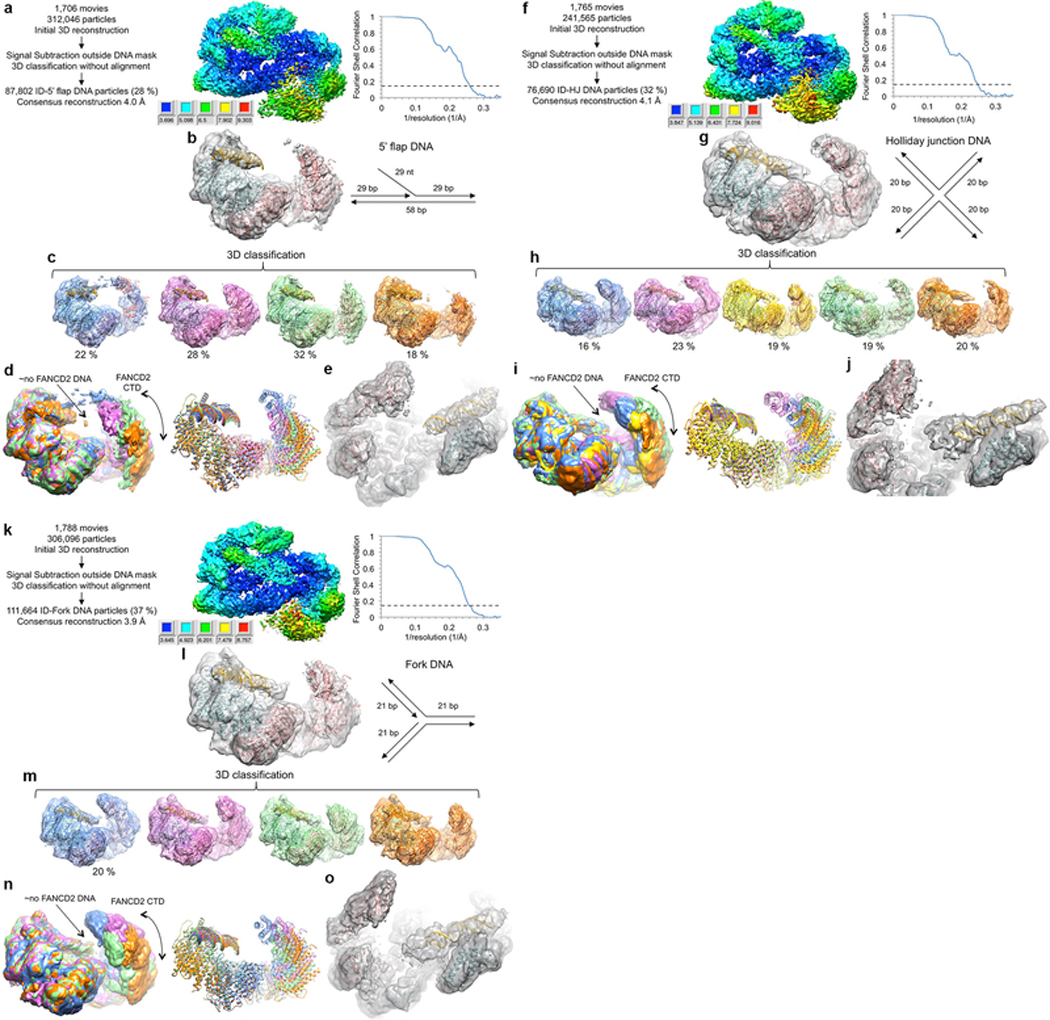
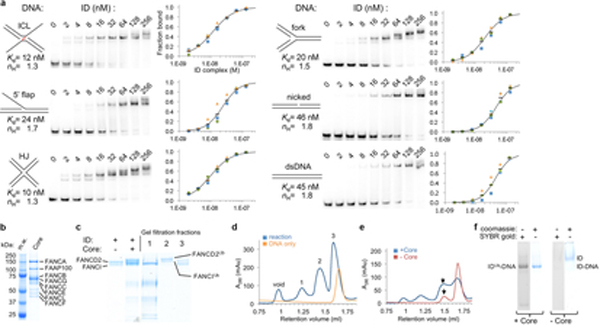
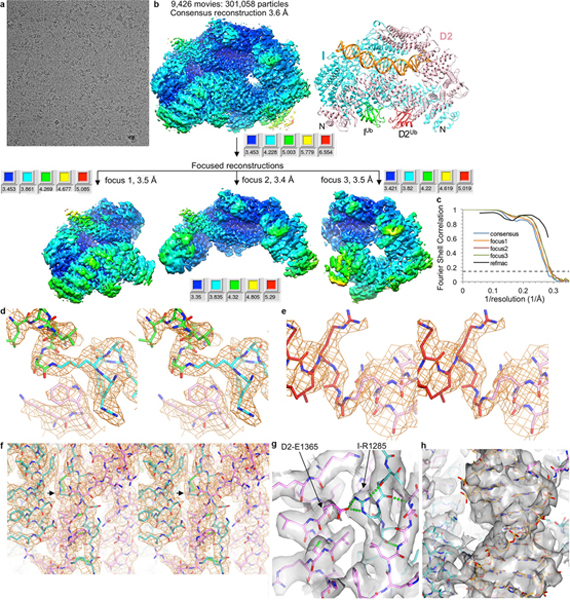
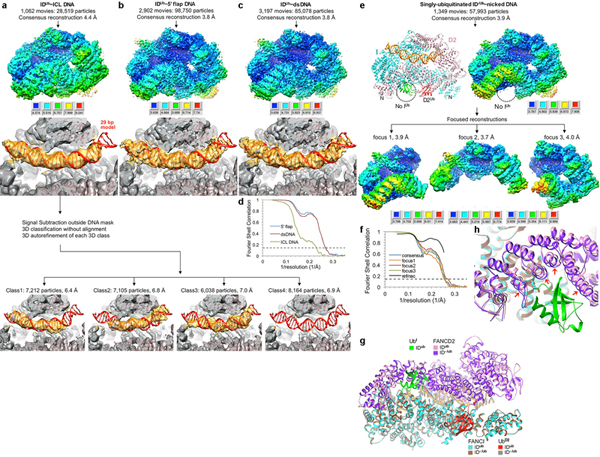
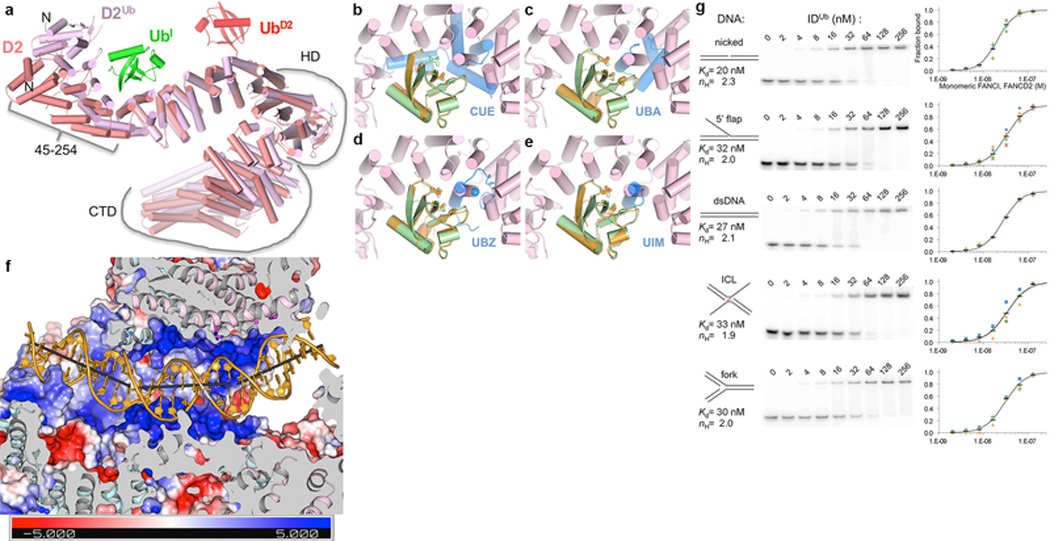
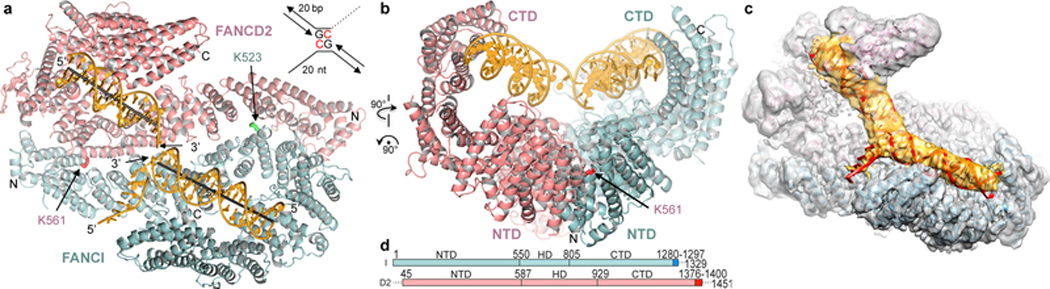
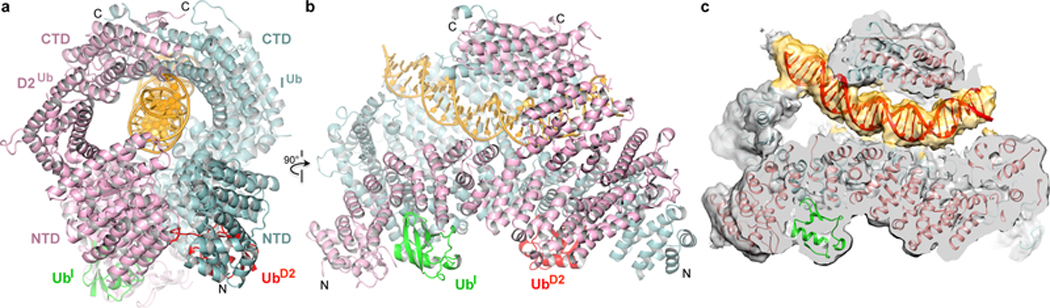
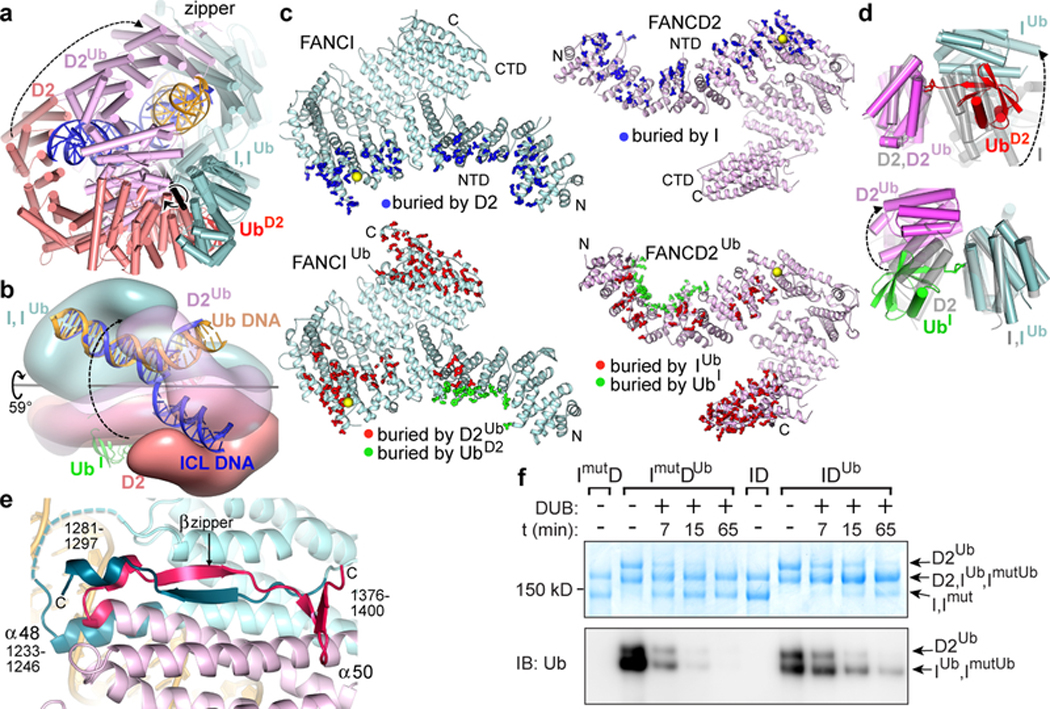
The ID complex, involving the proteins FANCI and FANCD2, is required for the repair of DNA interstrand crosslinks (ICL) and related lesions1. These proteins are mutated in Fanconi anaemia, a disease in which patients are predisposed to cancer. The Fanconi anaemia pathway of ICL repair is activated when a replication fork stalls at an ICL2; this triggers monoubiquitination of the ID complex, in which one ubiquitin molecule is conjugated to each of FANCI and FANCD2. Monoubiquitination of ID is essential for ICL repair by excision, translesion synthesis and homologous recombination; however, its function remains unknown1,3. Here we report a cryo-electron microscopy structure of the monoubiquitinated human ID complex bound to DNA, and reveal that it forms a closed ring that encircles the DNA. By comparison with the structure of the non-ubiquitinated ID complex bound to ICL DNA-which we also report here-we show that monoubiquitination triggers a complete rearrangement of the open, trough-like ID structure through the ubiquitin of one protomer binding to the other protomer in a reciprocal fashion. These structures-together with biochemical data-indicate that the monoubiquitinated ID complex loses its preference for ICL and related branched DNA structures, and becomes a sliding DNA clamp that can coordinate the subsequent repair reactions. Our findings also reveal how monoubiquitination in general can induce an alternative protein structure with a new function.
Figures
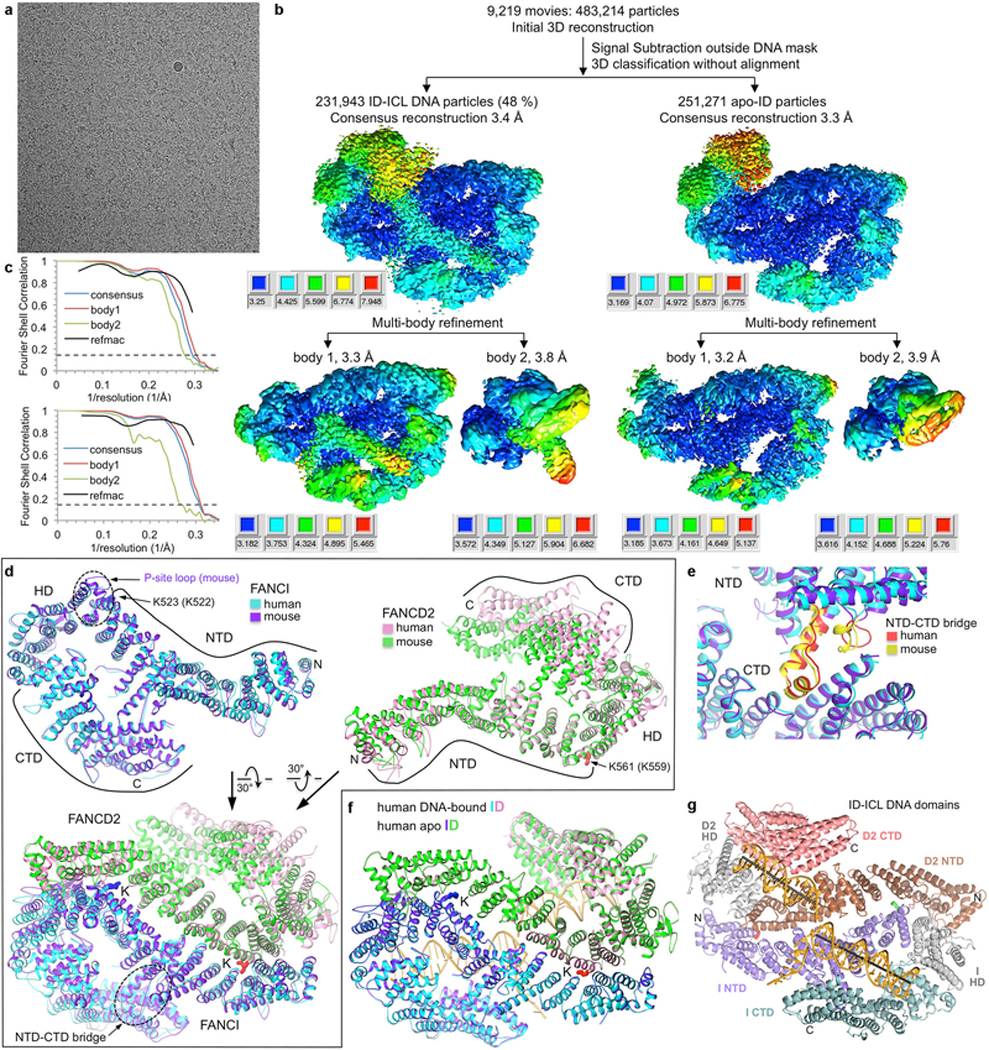
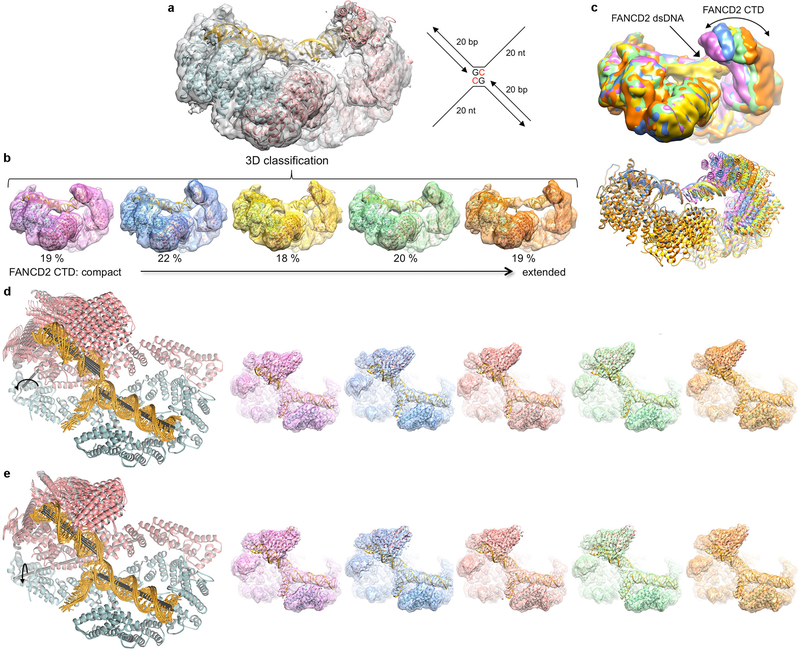
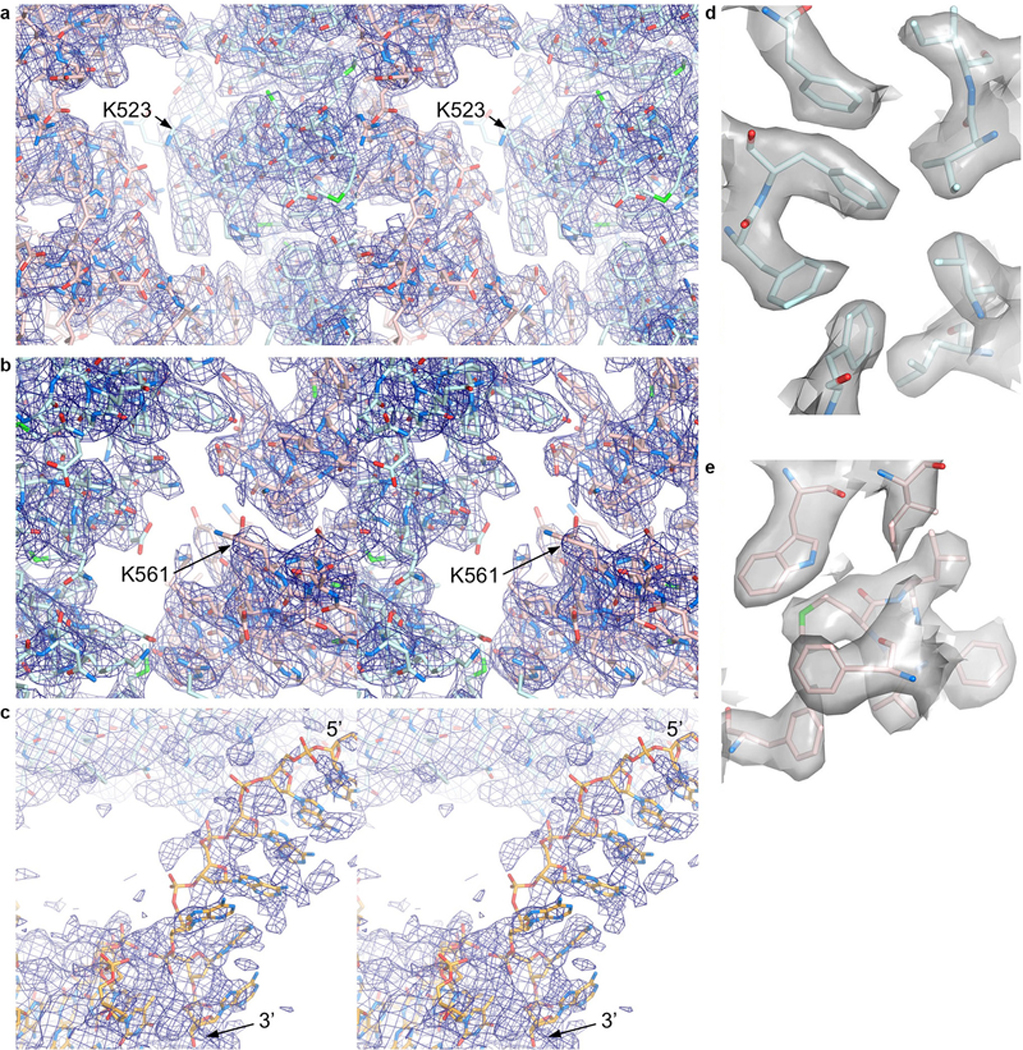
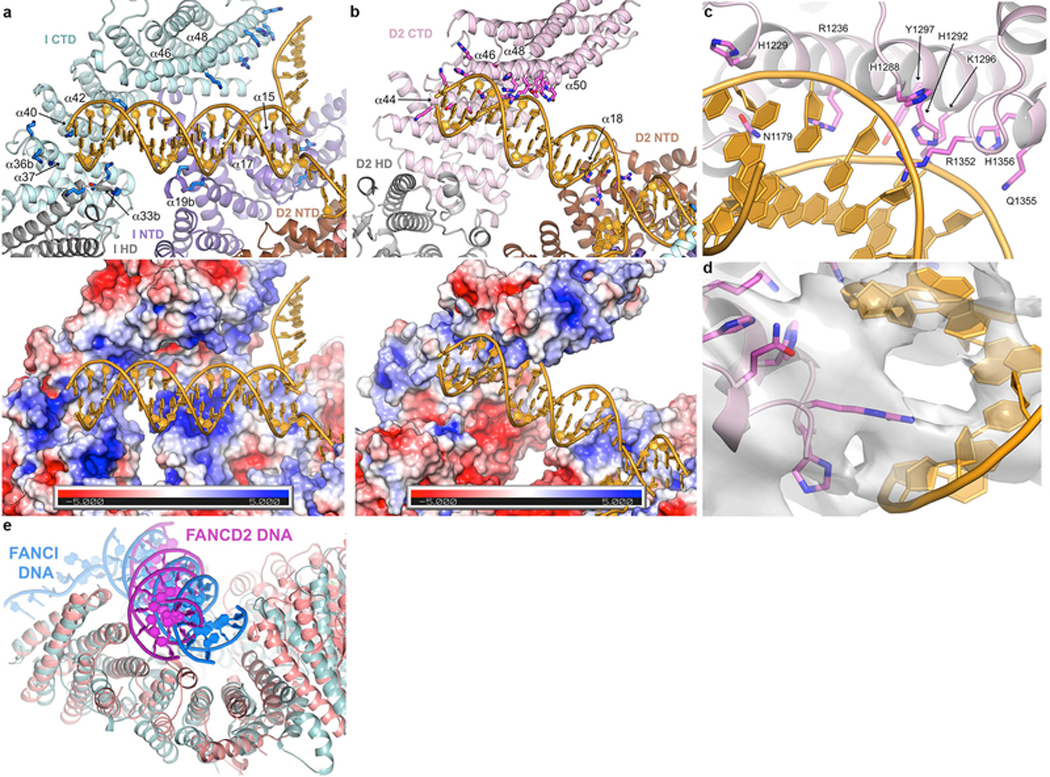
Comment in
-
Ubiquitination Causes Fanconi Anemia-Linked ID Complex Ring Formation.Cancer Discov. 2020 May;10(5):OF12. doi: 10.1158/2159-8290.CD-RW2020-043. Epub 2020 Mar 20. Cancer Discov. 2020. PMID: 32198134
-
Ubiquitilated Fanconi ID complex embraces DNA.Cell Res. 2020 Jul;30(7):554-555. doi: 10.1038/s41422-020-0345-2. Cell Res. 2020. PMID: 32472046 Free PMC article. No abstract available.
References
-
- Park WH et al. Direct DNA binding activity of the Fanconi anemia D2 protein. J Biol Chem 280, 23593–23598, doi:M503730200 [pii] 0.1074/jbc.M503730200 (2005). - PubMed
Extended Data References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
