

An intestinal zinc sensor regulates food intake and developmental growth
- PMID: 32269334
- PMCID: PMC8833092
- DOI: 10.1038/s41586-020-2111-5
An intestinal zinc sensor regulates food intake and developmental growth
Abstract
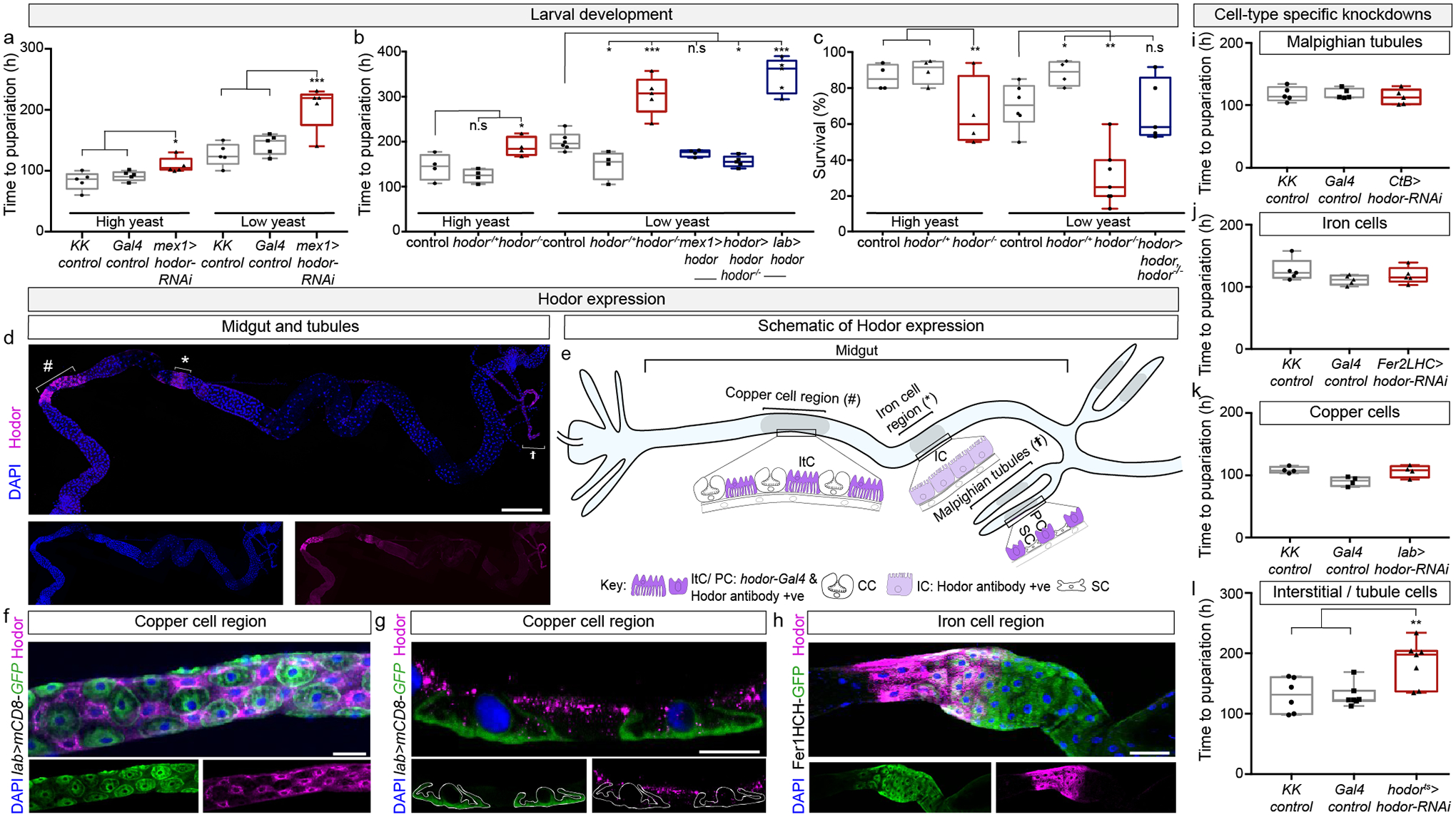
In cells, organs and whole organisms, nutrient sensing is key to maintaining homeostasis and adapting to a fluctuating environment1. In many animals, nutrient sensors are found within the enteroendocrine cells of the digestive system; however, less is known about nutrient sensing in their cellular siblings, the absorptive enterocytes1. Here we use a genetic screen in Drosophila melanogaster to identify Hodor, an ionotropic receptor in enterocytes that sustains larval development, particularly in nutrient-scarce conditions. Experiments in Xenopus oocytes and flies indicate that Hodor is a pH-sensitive, zinc-gated chloride channel that mediates a previously unrecognized dietary preference for zinc. Hodor controls systemic growth from a subset of enterocytes-interstitial cells-by promoting food intake and insulin/IGF signalling. Although Hodor sustains gut luminal acidity and restrains microbial loads, its effect on systemic growth results from the modulation of Tor signalling and lysosomal homeostasis within interstitial cells. Hodor-like genes are insect-specific, and may represent targets for the control of disease vectors. Indeed, CRISPR-Cas9 genome editing revealed that the single hodor orthologue in Anopheles gambiae is an essential gene. Our findings highlight the need to consider the instructive contributions of metals-and, more generally, micronutrients-to energy homeostasis.
Conflict of interest statement
COMPETING INTERESTS
The authors declare no competing interests.
Figures













Comment in
-
A zinc-sensing protein gives flies a gut feeling for growth.Nature. 2020 Apr;580(7802):187-188. doi: 10.1038/d41586-020-00728-8. Nature. 2020. PMID: 32203359 No abstract available.
References
-
- Colombani J et al. A nutrient sensor mechanism controls Drosophila growth. Cell 114, 739–749 (2003). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Other Literature Sources
Molecular Biology Databases