

Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function
- PMID: 32273340
- PMCID: PMC7242715
- DOI: 10.1074/jbc.RA120.012774
Single-molecule analysis reveals that regulatory light chains fine-tune skeletal myosin II function
Abstract
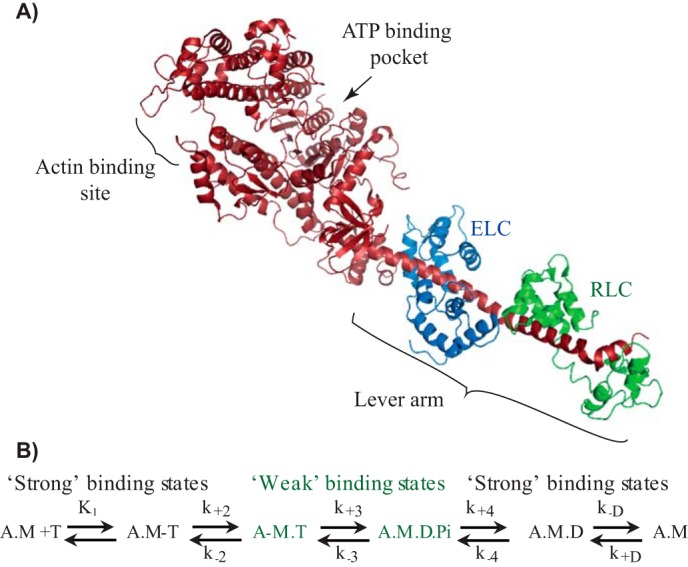
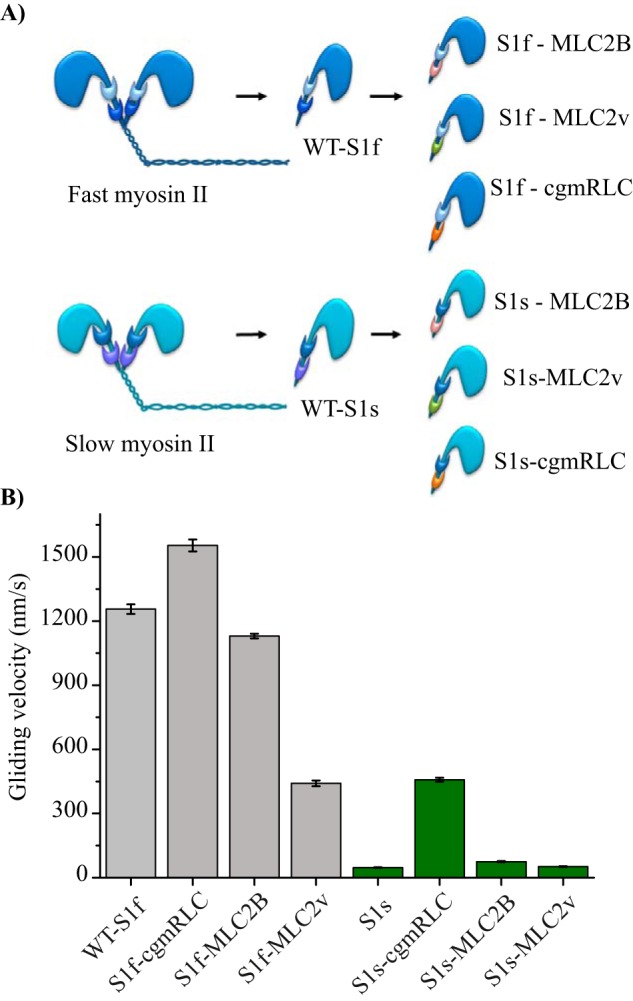
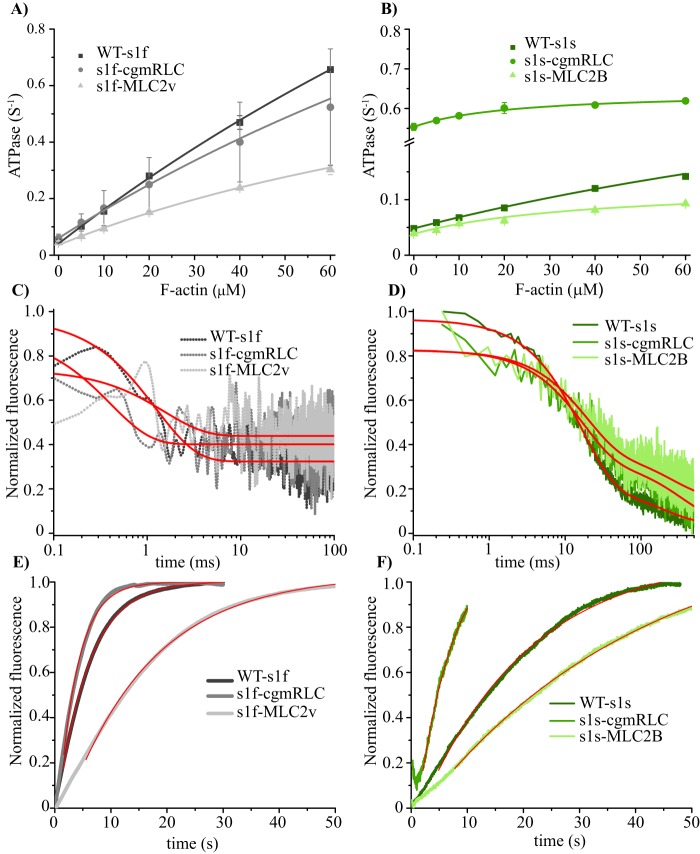
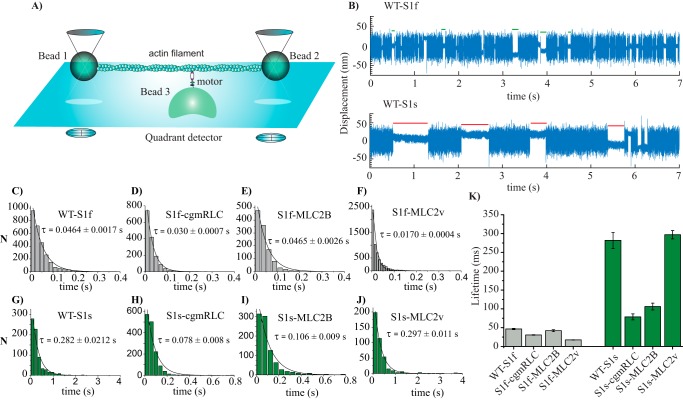
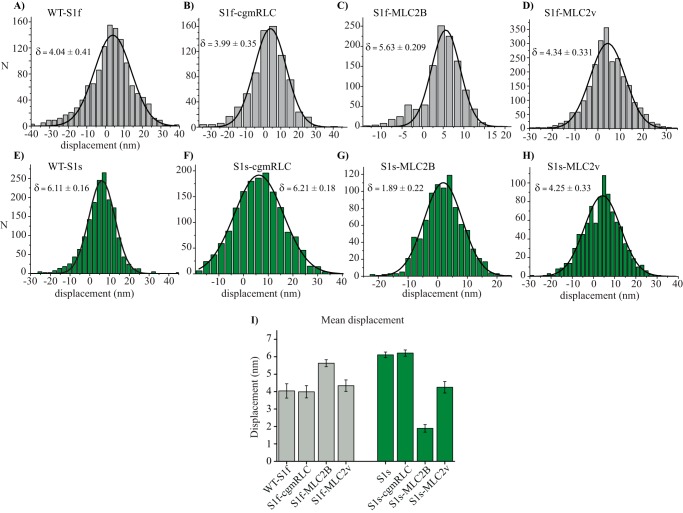
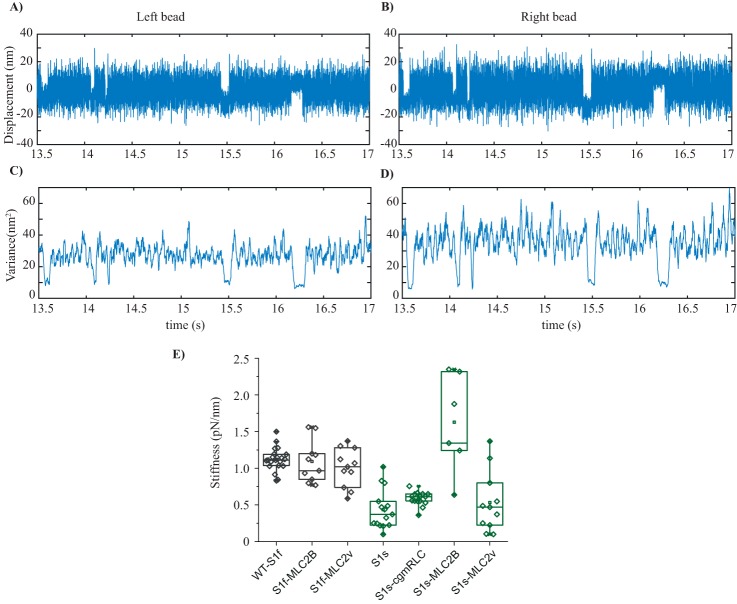
Myosin II is the main force-generating motor during muscle contraction. Myosin II exists as different isoforms that are involved in diverse physiological functions. One outstanding question is whether the myosin heavy chain (MHC) isoforms alone account for these distinct physiological properties. Unique sets of essential and regulatory light chains (RLCs) are known to assemble with specific MHCs, raising the intriguing possibility that light chains contribute to specialized myosin functions. Here, we asked whether different RLCs contribute to this functional diversification. To this end, we generated chimeric motors by reconstituting the MHC fast isoform (MyHC-IId) and slow isoform (MHC-I) with different light-chain variants. As a result of the RLC swapping, actin filament sliding velocity increased by ∼10-fold for the slow myosin and decreased by >3-fold for the fast myosin. Results from ensemble molecule solution kinetics and single-molecule optical trapping measurements provided in-depth insights into altered chemo-mechanical properties of the myosin motors that affect the sliding speed. Notably, we found that the mechanical output of both slow and fast myosins is sensitive to the RLC isoform. We therefore propose that RLCs are crucial for fine-tuning the myosin function.
Keywords: actin; actin filament gliding assay; molecular imaging; molecular motor; muscle contraction; myosin; myosin II; optical trapping; optical tweezers; regulatory light chain; single-molecule analysis; single-molecule biophysics; solution kinetics.
© 2020 Nayak et al.
Conflict of interest statement
The authors declare that they have no conflicts of interest with the contents of this article
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
