

MicroRNAs as Emerging Regulators of Signaling in the Tumor Microenvironment
- PMID: 32276464
- PMCID: PMC7225969
- DOI: 10.3390/cancers12040911
MicroRNAs as Emerging Regulators of Signaling in the Tumor Microenvironment
Abstract
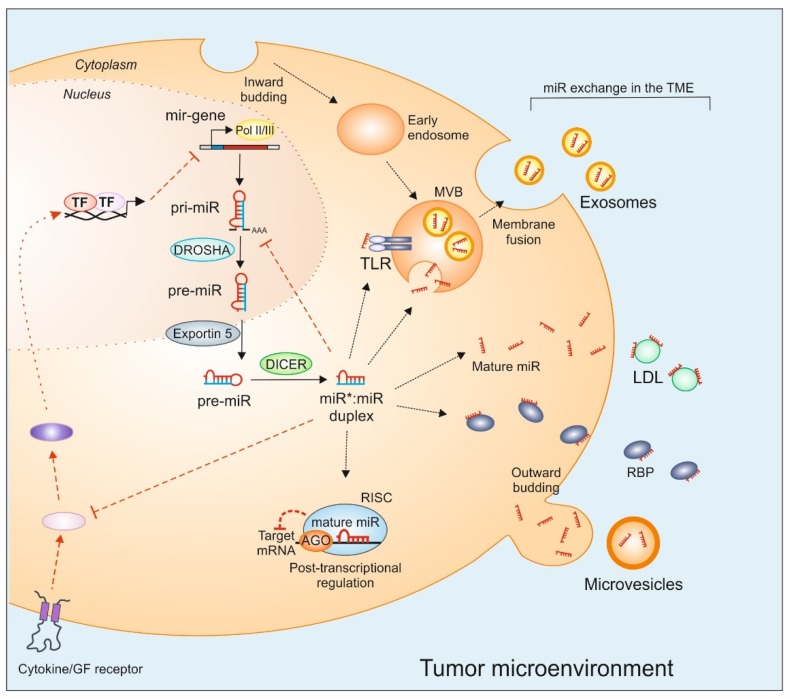
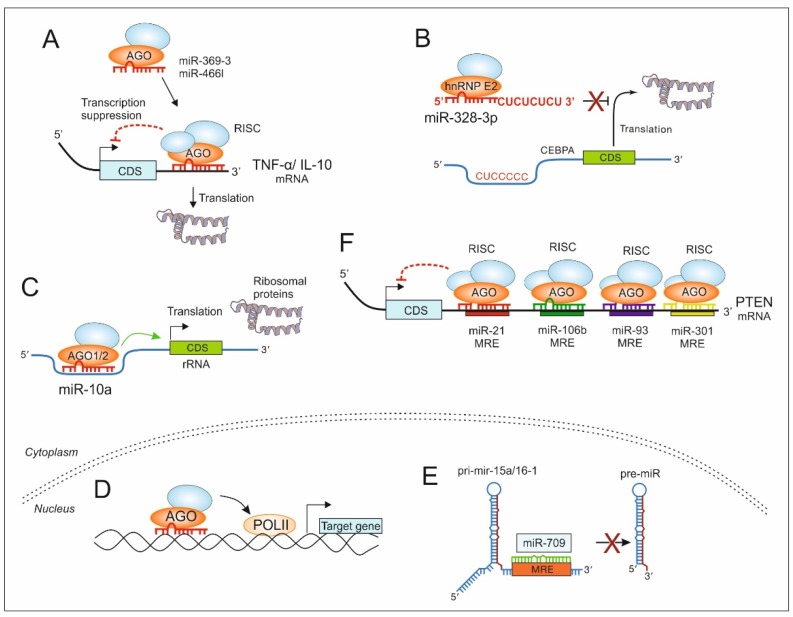
A myriad of signaling molecules in a heuristic network of the tumor microenvironment (TME) pose a challenge and an opportunity for novel therapeutic target identification in human cancers. MicroRNAs (miRs), due to their ability to affect signaling pathways at various levels, take a prominent space in the quest of novel cancer therapeutics. The role of miRs in cancer initiation, progression, as well as in chemoresistance, is being increasingly investigated. The canonical function of miRs is to target mRNAs for post-transcriptional gene silencing, which has a great implication in first-order regulation of signaling pathways. However, several reports suggest that miRs also perform non-canonical functions, partly due to their characteristic non-coding small RNA nature. Examples emerge when they act as ligands for toll-like receptors or perform second-order functions, e.g., to regulate protein translation and interactions. This review is a compendium of recent advancements in understanding the role of miRs in cancer signaling and focuses on the role of miRs as novel regulators of the signaling pathway in the TME.
Keywords: RNA therapeutics; breast cancer; cancer; carcinoma; inflammation; microRNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Odes E.J., Randolph-Quinney P.S., Steyn M., Throckmorton Z., Smilg J.S., Zipfel B., Augustine T.N., Beer F.d., Hoffman J.W., Franklin R.D., et al. Earliest hominin cancer: 1.7-million-year-old osteosarcoma from Swartkrans Cave, South Africa. S. Afr. J. Sci. 2016;112 doi: 10.17159/sajs.2016/20150471. - DOI
-
- Randolph-Quinney P.S., Williams S.A., Steyn M., Meyer M.R., Smilg J.S., Churchill S.E., Odes E.J., Augustine T., Tafforeau P., Berger L.R. Osteogenic tumour in Australopithecus sediba: Earliest hominin evidence for neoplastic disease. S. Afr. J. Sci. 2016;112 doi: 10.17159/sajs.2016/20150470. - DOI
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
