

MCPH1 Lack of Function Enhances Mitotic Cell Sensitivity Caused by Catalytic Inhibitors of Topo II
- PMID: 32276518
- PMCID: PMC7231051
- DOI: 10.3390/genes11040406
MCPH1 Lack of Function Enhances Mitotic Cell Sensitivity Caused by Catalytic Inhibitors of Topo II
Abstract
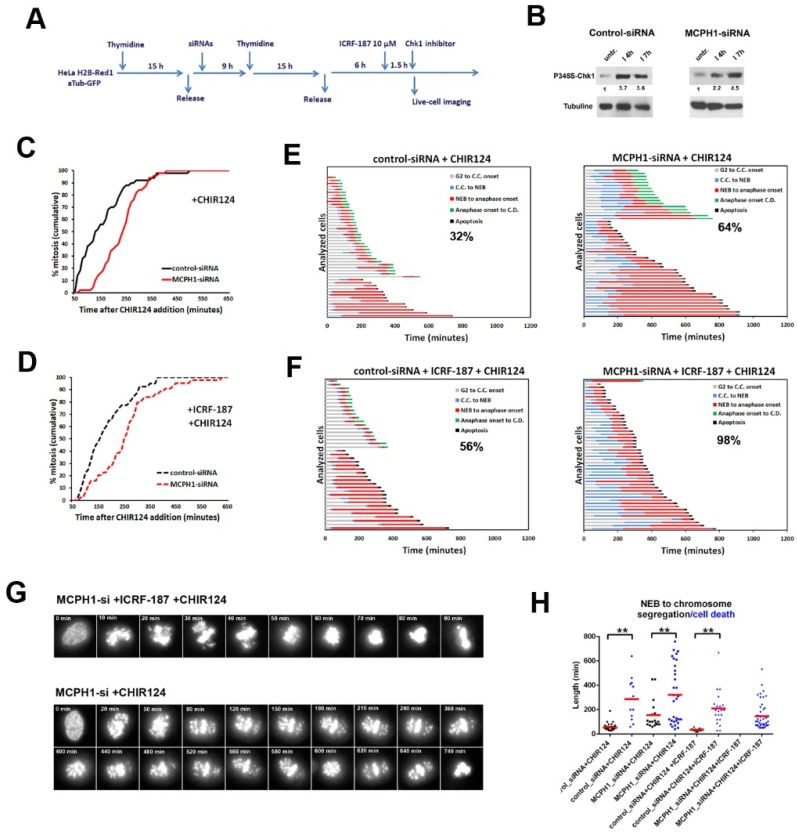
The capacity of Topoisomerase II (Topo II) to remove DNA catenations that arise after replication is essential to ensure faithful chromosome segregation. Topo II activity is monitored during G2 by a specific checkpoint pathway that delays entry into mitosis until the chromosomes are properly decatenated. Recently, we demonstrated that the mitotic defects that are characteristic of cells depleted of MCPH1 function, a protein mutated in primary microcephaly, are not a consequence of a weakened G2 decatenation checkpoint response. However, the mitotic defects could be accounted for by a minor defect in the activity of Topo II during G2/M. To test this hypothesis, we have tracked at live single cell resolution the dynamics of mitosis in MCPH1 depleted HeLa cells upon catalytic inhibition of Topo II. Our analyses demonstrate that neither chromosome alignment nor segregation are more susceptible to minor perturbation in decatenation in MCPH1 deficient cells, as compared with control cells. Interestingly, MCPH1 depleted cells were more prone to mitotic cell death when decatenation was perturbed. Furthermore, when the G2 arrest that was induced by catalytic inhibition of Topo II was abrogated by Chk1 inhibition, the incidence of mitotic cell death was also increased. Taken together, our data suggest that the MCPH1 lack of function increases mitotic cell hypersensitivity to the catalytic inhibition of Topo II.
Keywords: ICRF; MCPH1; Topoisomerase II; anaphase errors; chromosome condensation; chromosome segregation; decatenation checkpoint; mitotic catastrophe; mitotic cell death.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
Figures



References
-
- Giménez-Abián J.F., Clarke D.J., Giménez-Martín G., Weingartner M., Giménez-Abián M.I., Carballo J.A., Díaz de la Espina S.M., Bögre L., De la Torre C. DNA catenations that link sister chromatids until the onset of anaphase are maintained by a checkpoint mechanism. Eur. J. Cell Biol. 2002;81:9–16. doi: 10.1078/0171-9335-00226. - DOI - PubMed
-
- Clarke D.J., Johnson R.T., Downes C.S. Topoisomerase II inhibition prevents anaphase chromatid segregation in mammalian cells independently of the generation of DNA strand breaks. J. Cell Sci. 1993;569:563–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
