

The MID1 gene product in physiology and disease
- PMID: 32283114
- PMCID: PMC8011326
- DOI: 10.1016/j.gene.2020.144655
The MID1 gene product in physiology and disease
Abstract
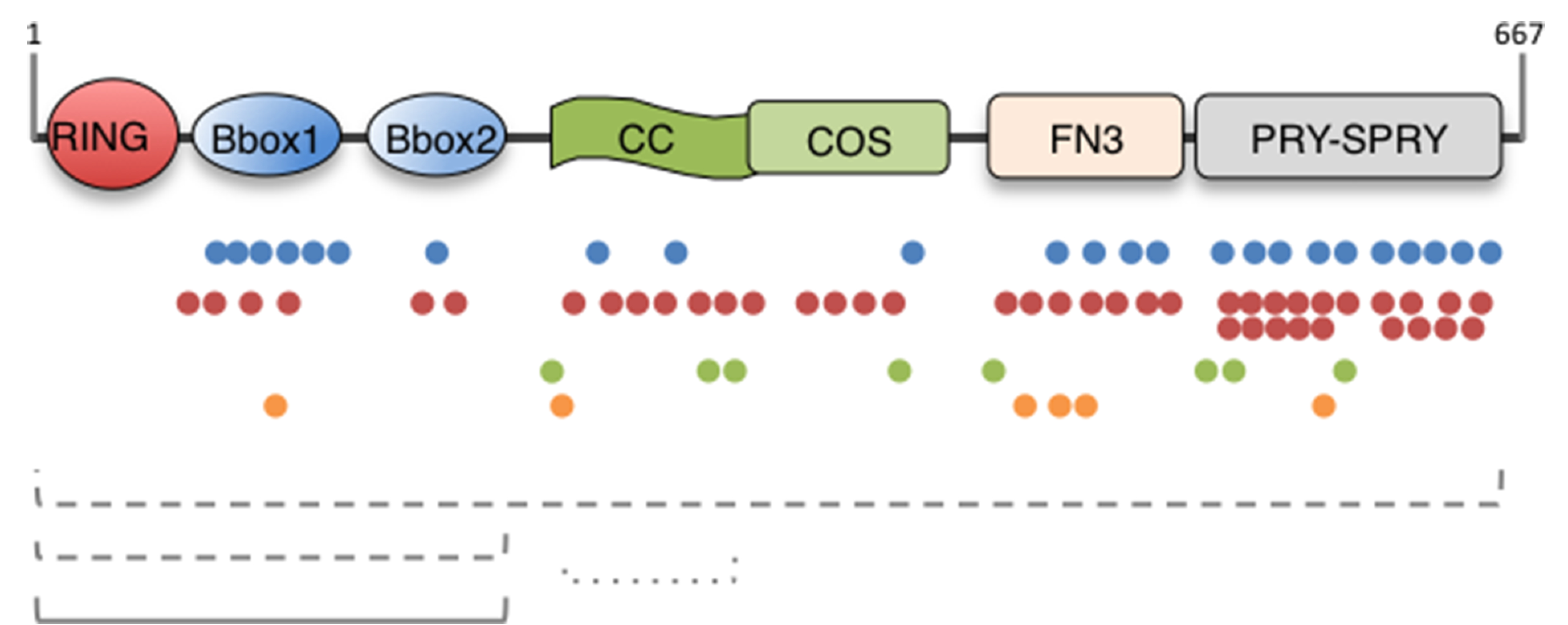
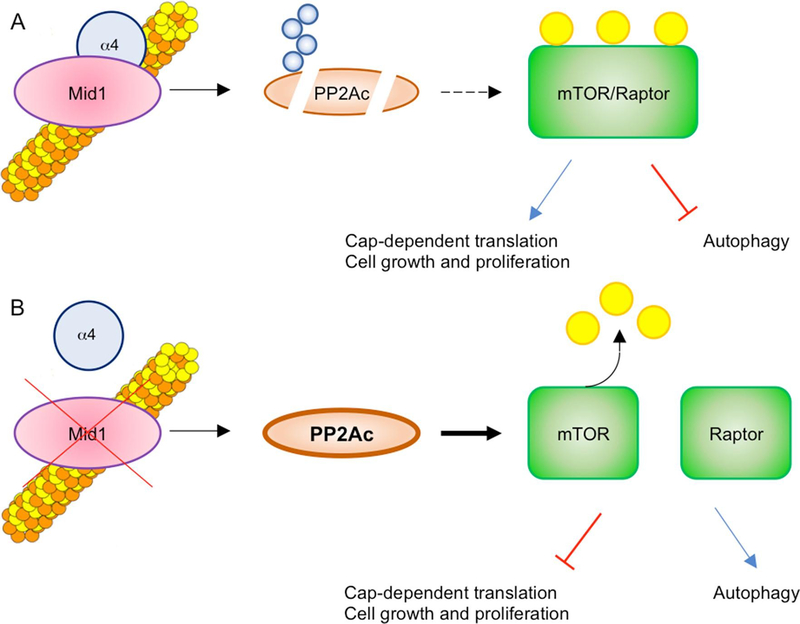
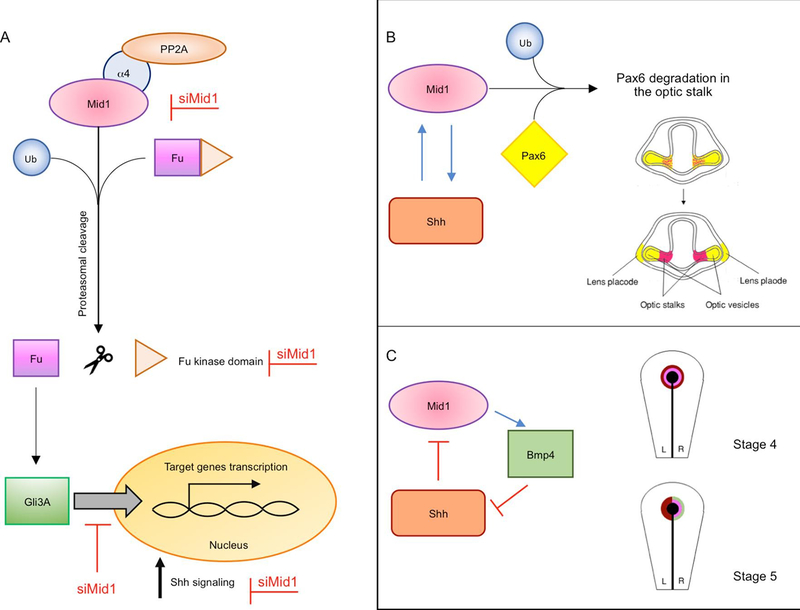
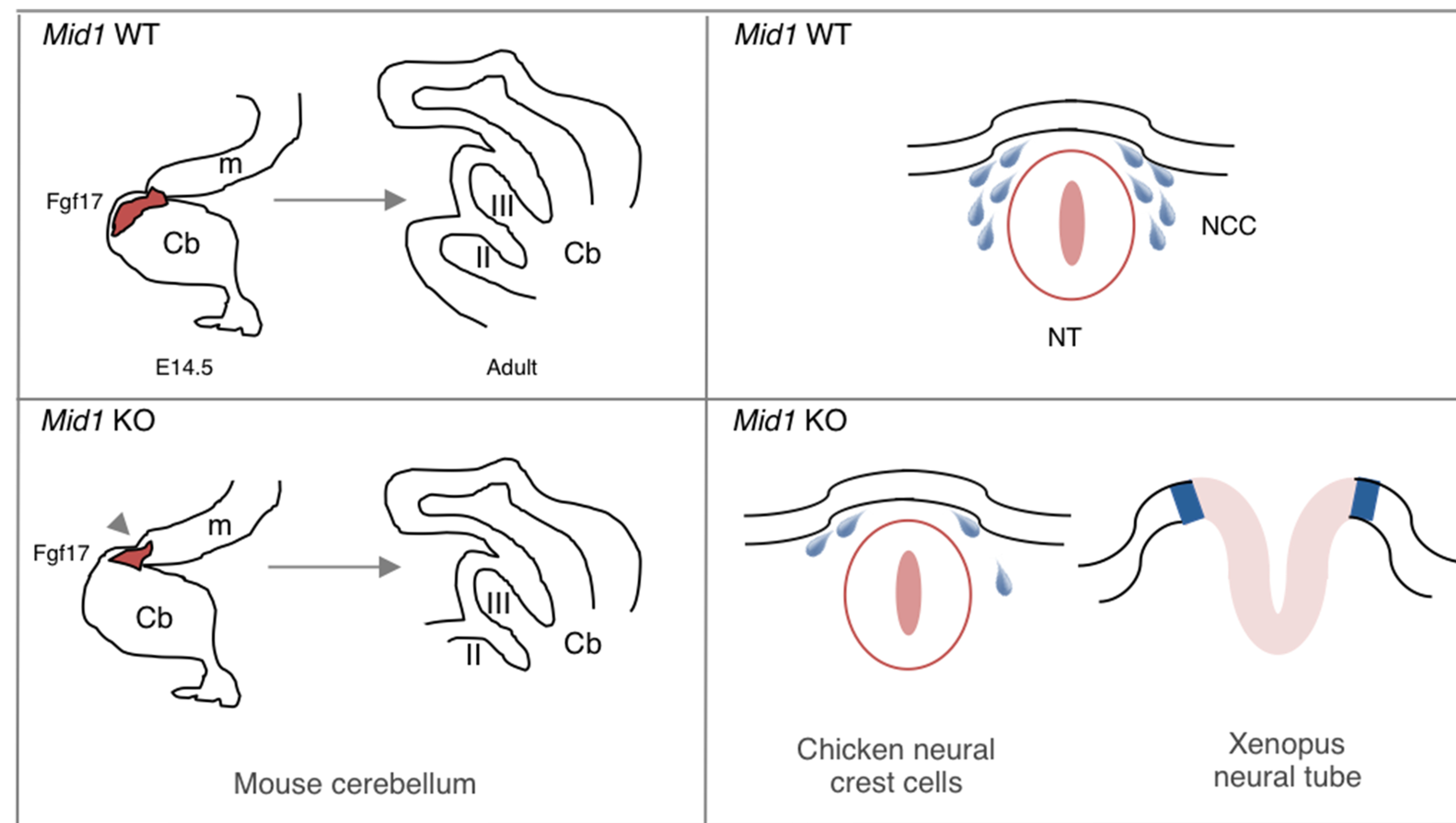
MID1 is an E3 ubiquitin ligase of the Tripartite Motif (TRIM) subfamily of RING-containing proteins, hence also known as TRIM18. MID1 is a microtubule-binding protein found in complex with the catalytic subunit of PP2A (PP2Ac) and its regulatory subunit alpha 4 (α4). To date, several substrates and interactors of MID1 have been described, providing evidence for the involvement of MID1 in a plethora of essential biological processes, especially during embryonic development. Mutations in the MID1 gene are responsible of the X-linked form of Opitz syndrome (XLOS), a multiple congenital disease characterised by defects in the development of midline structures during embryogenesis. Here, we review MID1-related physiological mechanisms as well as the pathological implication of the MID1 gene in XLOS and in other clinical conditions.
Copyright © 2020 Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of Competing Interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
References
-
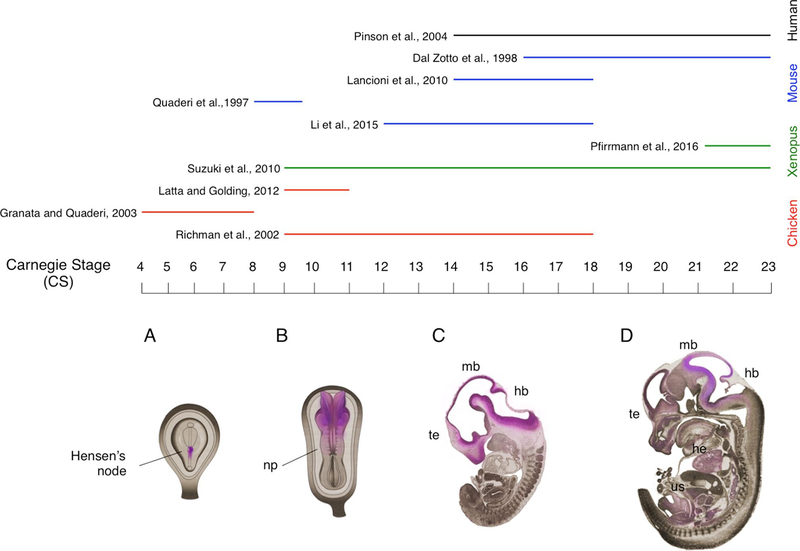
- Abramyan J, Richman JM, 2018. Craniofacial development: discoveries made in the chicken embryo. Int. J. Dev. Biol 62, 97–107. - PubMed
-
- Aranda-Orgilles B, Rutschow D, Zeller R, Karagiannidis AI, Kohler A, Chen C, Wilson T, Krause S, Roepcke S, Lilley D, et al., 2011. Protein phosphatase 2A (PP2A)-specific ubiquitin ligase MID1 is a sequence-dependent regulator of translation efficiency controlling 3-phosphoinositide-dependent protein kinase-1 (PDPK-1). J. Biol. Chem 286, 39945–39957. - PMC - PubMed
-
- Arigoni M, Barutello G, Riccardo F, Ercole E, Cantarella D, Orso F, Conti L, Lanzardo S, Taverna D, Merighi I, et al., 2013. miR-135b coordinates progression of ErbB2-driven mammary carcinomas through suppression of MID1 and MTCH2. Am. J. Pathol 182, 2058–2070. - PubMed
