

Retinoic Acid Modulates Hyperactive T Cell Responses and Protects Vitamin A-Deficient Mice against Persistent Lymphocytic Choriomeningitis Virus Infection
- PMID: 32284332
- PMCID: PMC7371257
- DOI: 10.4049/jimmunol.1901091
Retinoic Acid Modulates Hyperactive T Cell Responses and Protects Vitamin A-Deficient Mice against Persistent Lymphocytic Choriomeningitis Virus Infection
Abstract
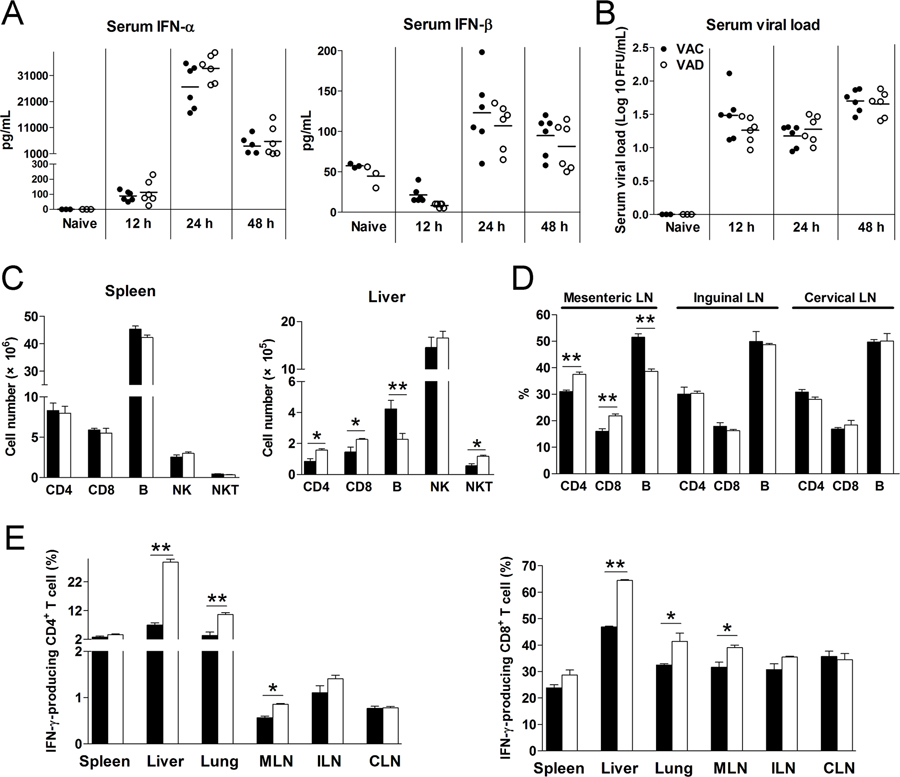
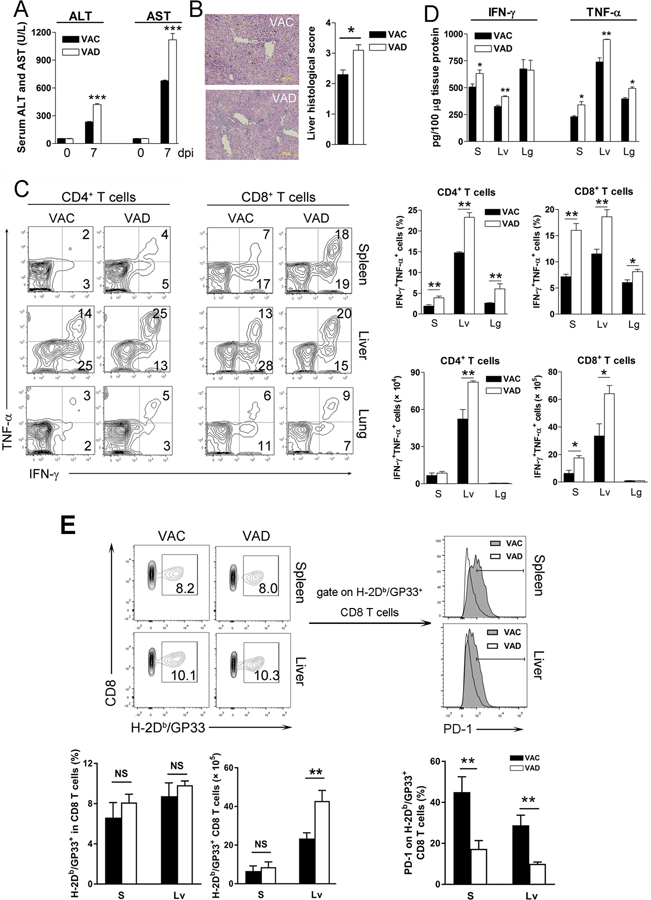
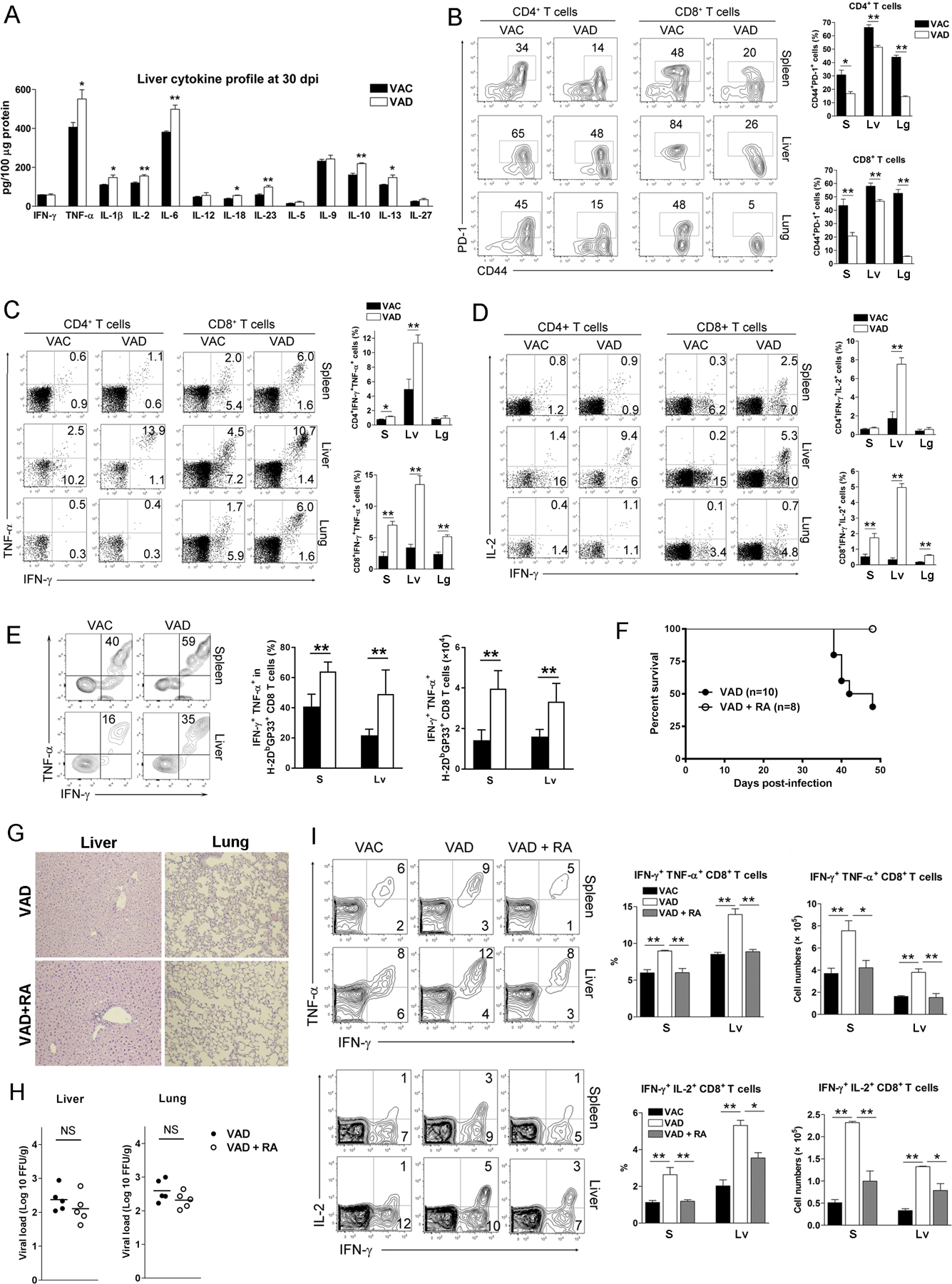
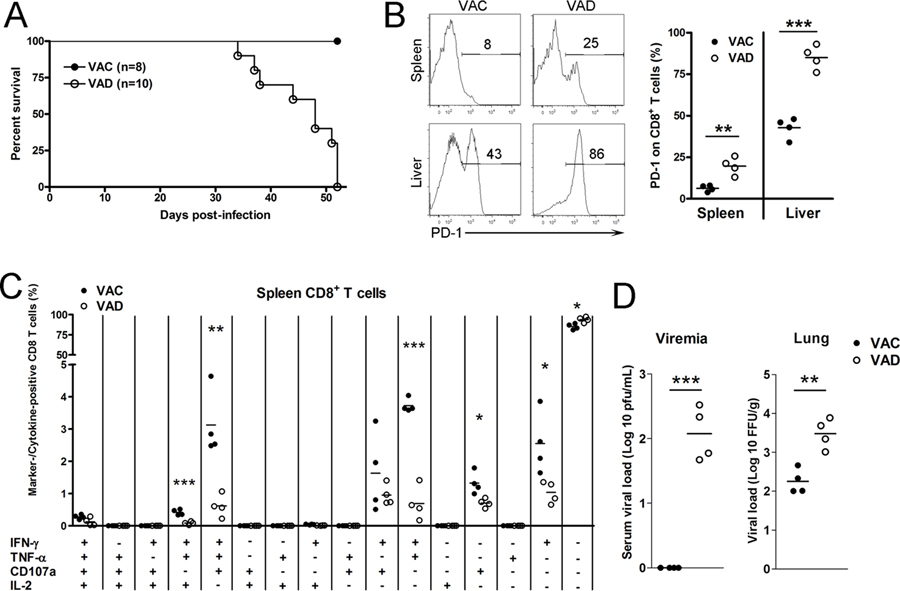
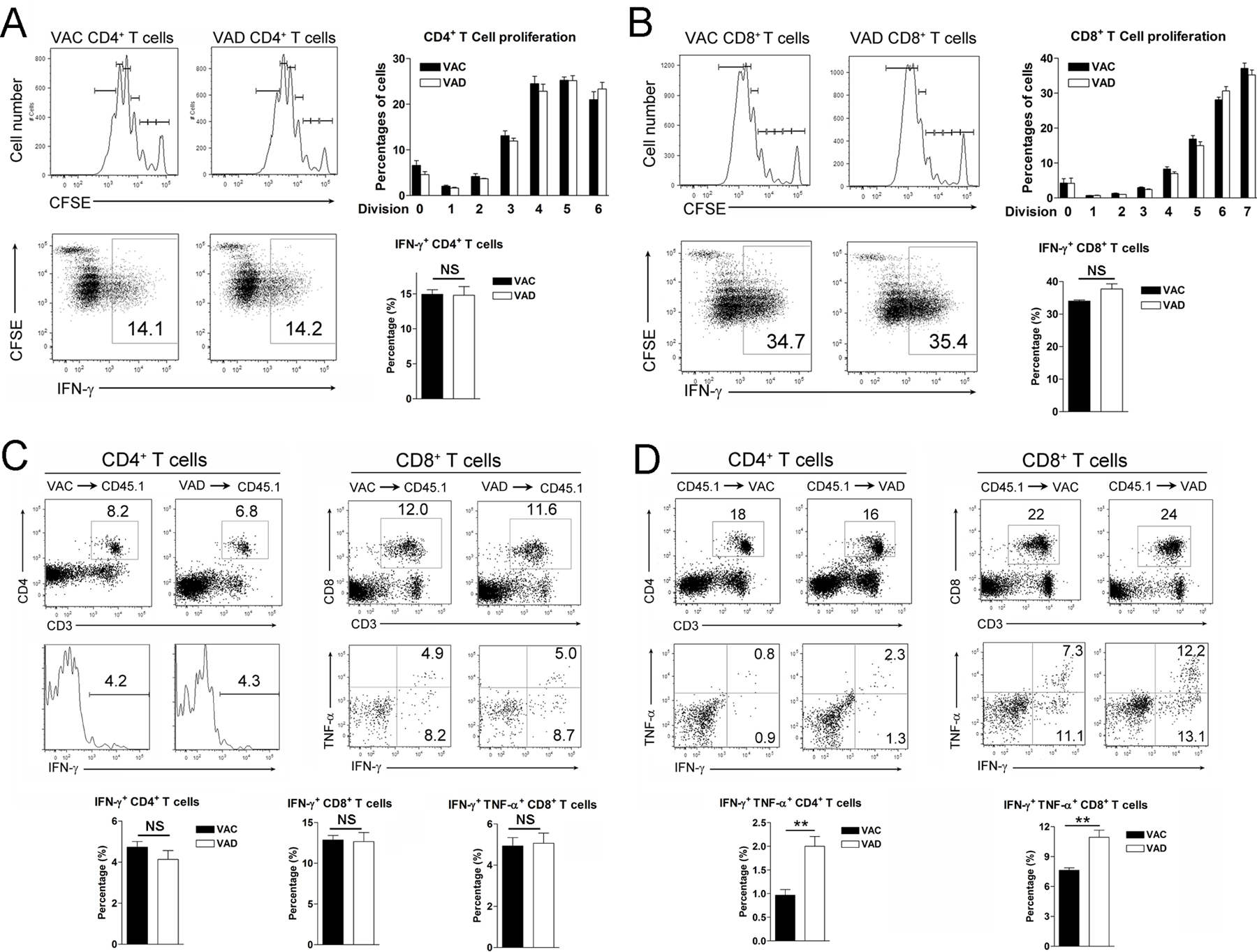
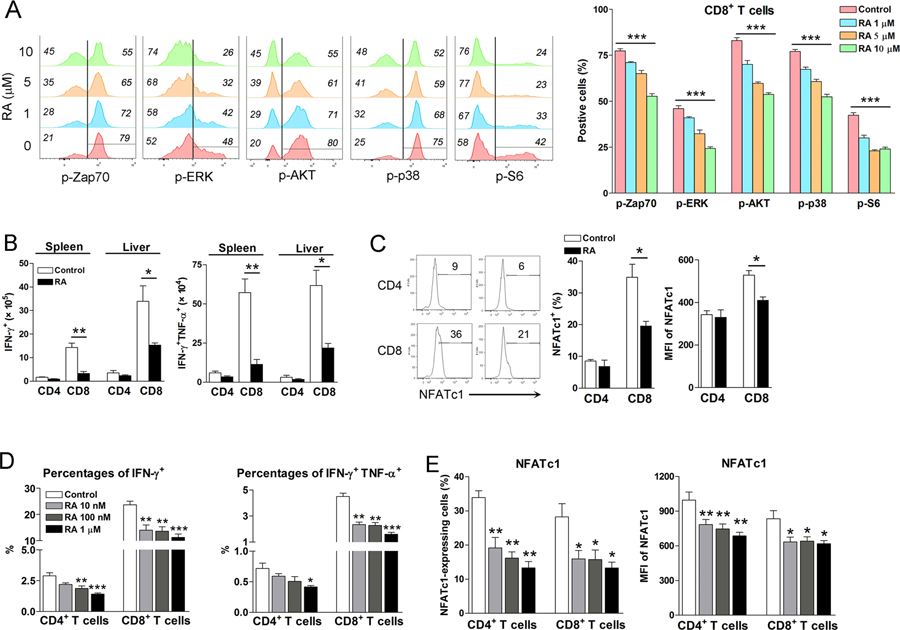
Vitamin A deficiency (VAD) is a major public health problem and is associated with increased host susceptibility to infection; however, how VAD influences viral infection remains unclear. Using a persistent lymphocytic choriomeningitis virus infection model, we showed in this study that although VAD did not alter innate type I IFN production, infected VAD mice had hyperactive, virus-specific T cell responses at both the acute and contraction stages, showing significantly decreased PD-1 but increased cytokine (IFN-γ, TNF-α, and IL-2) expression by T cells. Compared with control mice, VAD mice displayed excessive inflammation and more severe liver pathology, with increased death during persistent infection. Of note, supplements of all-trans retinoic acid (RA), one of the important metabolites of vitamin A, downregulated hyperactive T cell responses and rescued the persistently infected VAD mice. By using adoptive transfer of splenocytes, we found that the environmental vitamin A or its metabolites acted as rheostats modulating antiviral T cells. The analyses of T cell transcriptional factors and signaling pathways revealed the possible mechanisms of RA, as its supplements inhibited the abundance of NFATc1 (NFAT 1), a key regulator for T cell activation. Also, following CD3/CD28 cross-linking stimulation, RA negatively regulated the TCR-proximal signaling in T cells, via decreased phosphorylation of Zap70 and its downstream signals, including phosphorylated AKT, p38, ERK, and S6, respectively. Together, our data reveal VAD-mediated alterations in antiviral T cell responses and highlight the potential utility of RA for modulating excessive immune responses and tissue injury in infectious diseases.
Copyright © 2020 by The American Association of Immunologists, Inc.
Conflict of interest statement
Disclosures: The authors declare no financial or commercial conflict of interest.
Figures
References
-
- Stevens GA, Bennett JE, Hennocq Q, Lu Y, De-Regil LM, Rogers L, Danaei G, Li G, White RA, Flaxman SR, Oehrle SP, Finucane MM, Guerrero R, Bhutta ZA, Then-Paulino A, Fawzi W, Black RE, and Ezzati M 2015. Trends and mortality effects of vitamin A deficiency in children in 138 low-income and middle-income countries between 1991 and 2013: a pooled analysis of population-based surveys. Lancet Glob Health 3:e528–536. - PubMed
-
- Baum M, Cassetti L, Bonvehi P, Shor-Posner G, Lu Y, and Sauberlich H 1994. Inadequate dietary intake and altered nutrition status in early HIV-1 infection. Nutrition 10:16–20. - PubMed
-
- Semba RD, Caiaffa WT, Graham NM, Cohn S, and Vlahov D 1995. Vitamin A deficiency and wasting as predictors of mortality in human immunodeficiency virus-infected injection drug users. J Infect Dis 171:1196–1202. - PubMed
-
- Peres WA, Chaves GV, Goncalves JC, Ramalho A, and Coelho HS 2011. Vitamin A deficiency in patients with hepatitis C virus-related chronic liver disease. Br J Nutr 106:1724–1731. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
