

Soft extracellular matrix enhances inflammatory activation of mesenchymal stromal cells to induce monocyte production and trafficking
- PMID: 32284989
- PMCID: PMC7141831
- DOI: 10.1126/sciadv.aaw0158
Soft extracellular matrix enhances inflammatory activation of mesenchymal stromal cells to induce monocyte production and trafficking
Abstract
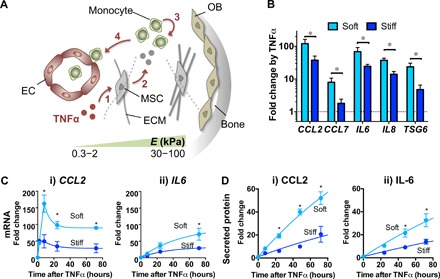
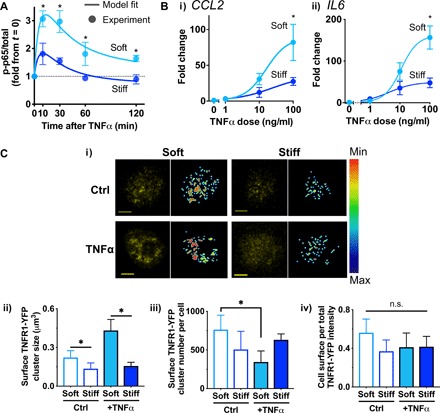
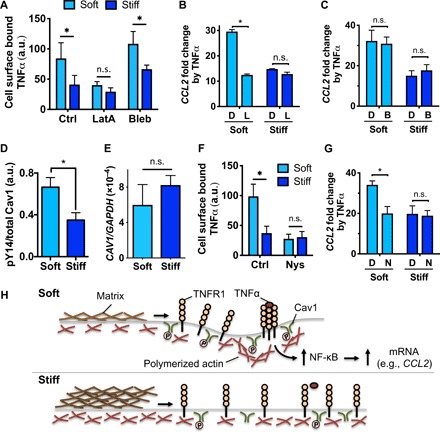
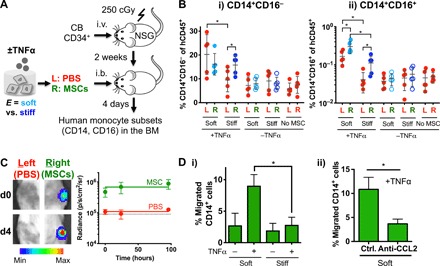
Mesenchymal stromal cells (MSCs) modulate immune cells to ameliorate multiple inflammatory pathologies. Biophysical signals that regulate this process are poorly defined. By engineering hydrogels with tunable biophysical parameters relevant to bone marrow where MSCs naturally reside, we show that soft extracellular matrix maximizes the ability of MSCs to produce paracrine factors that have been implicated in monocyte production and chemotaxis upon inflammatory stimulation by tumor necrosis factor-α (TNFα). Soft matrix increases clustering of TNF receptors, thereby enhancing NF-κB activation and downstream gene expression. Actin polymerization and lipid rafts, but not myosin-II contractility, regulate mechanosensitive activation of MSCs by TNFα. We functionally demonstrate that human MSCs primed with TNFα in soft matrix enhance production of human monocytes in marrow of xenografted mice and increase trafficking of monocytes via CCL2. The results suggest the importance of biophysical signaling in tuning inflammatory activation of stromal cells to control the innate immune system.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works. Distributed under a Creative Commons Attribution NonCommercial License 4.0 (CC BY-NC).
Figures
References
-
- Yue R., Zhou B. O., Shimada I. S., Zhao Z., Morrison S. J., Leptin receptor promotes adipogenesis and reduces osteogenesis by regulating mesenchymal stromal cells in adult bone marrow. Cell Stem Cell 18, 782–796 (2016). - PubMed
-
- Bartholomew A., Sturgeon C., Siatskas M., Ferrer K., McIntosh K., Patil S., Hardy W., Devine S., Ucker D., Deans R., Moseley A., Hoffman R., Mesenchymal stem cells suppress lymphocyte proliferation in vitro and prolong skin graft survival in vivo. Exp. Hematol. 30, 42–48 (2002). - PubMed
-
- Le Blanc K., Mougiakakos D., Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immunol. 12, 383–396 (2012). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
