

Eliminating effects of particle adsorption to the air/water interface in single-particle cryo-electron microscopy: Bacterial RNA polymerase and CHAPSO
- PMID: 32285040
- PMCID: PMC7153306
- DOI: 10.1016/j.yjsbx.2019.100005
Eliminating effects of particle adsorption to the air/water interface in single-particle cryo-electron microscopy: Bacterial RNA polymerase and CHAPSO
Abstract
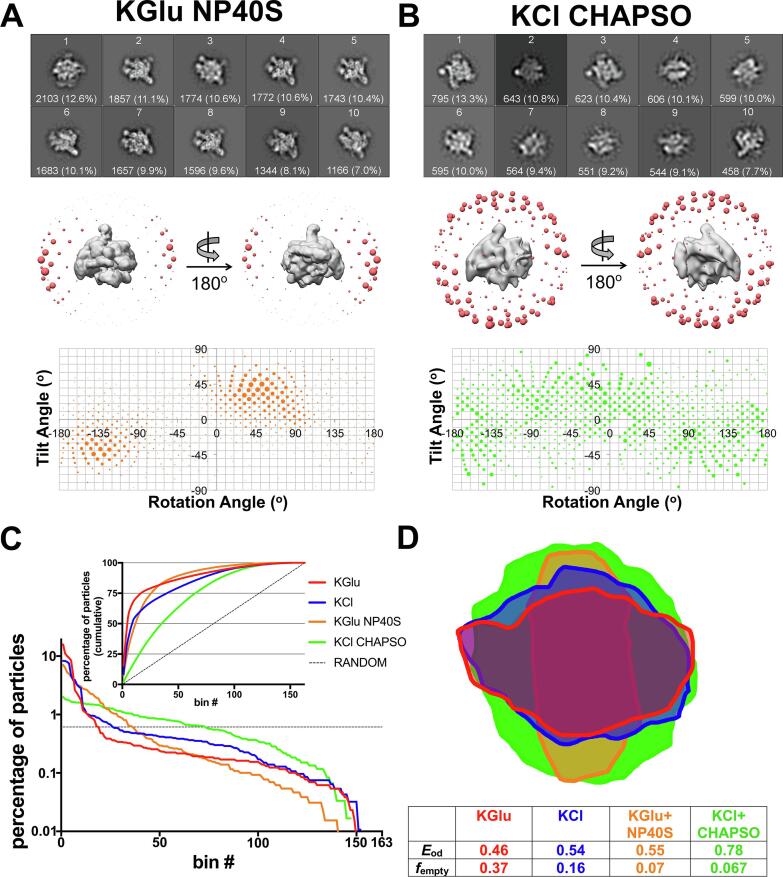
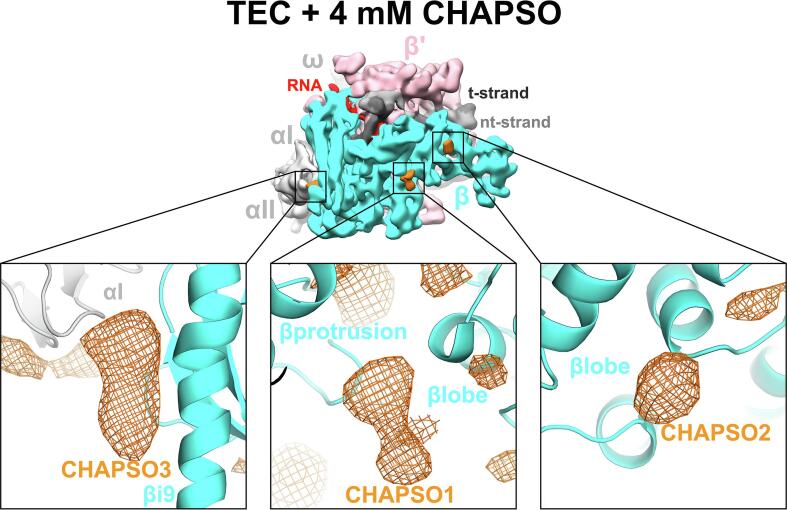
Preferred particle orientation presents a major challenge for many single particle cryo-electron microscopy (cryo-EM) samples. Orientation bias limits the angular information used to generate three-dimensional maps and thus affects the reliability and interpretability of the structural models. The primary cause of preferred orientation is presumed to be due to adsorption of the particles at the air/water interface during cryo-EM grid preparation. To ameliorate this problem, detergents are often added to cryo-EM samples to alter the properties of the air/water interface. We have found that many bacterial transcription complexes suffer severe orientation bias when examined by cryo-EM. The addition of non-ionic detergents, such as NP-40, does not remove the orientation bias but the Zwitter-ionic detergent CHAPSO significantly broadens the particle orientation distributions, yielding isotropically uniform maps. We used cryo-electron tomography to examine the particle distribution within the ice layer of cryo-EM grid preparations of Escherichia coli 6S RNA/RNA polymerase holoenzyme particles. In the absence of CHAPSO, essentially all of the particles are located at the ice surfaces. CHAPSO at the critical micelle concentration eliminates particle absorption at the air/water interface and allows particles to randomly orient in the vitreous ice layer. We find that CHAPSO eliminates orientation bias for a wide range of bacterial transcription complexes containing E. coli or Mycobacterium tuberculosis RNA polymerases. Findings of this study confirm the presumed basis for how detergents can help remove orientation bias in cryo-EM samples and establishes CHAPSO as a useful tool to facilitate cryo-EM studies of bacterial transcription complexes.
Keywords: Cryo-electron tomography.
Conflict of interest statement
Declarations of interest: None
Figures






References
-
- Bae B., Davis E., Brown D., Campbell E.A., Wigneshweraraj S.R., Darst S.A. Phage T7 Gp2 inhibition of Escherichia coli RNA polymerase involves misappropriation of σ70 domain 1.1. Proceedings of the National Academy of Sciences. 2013;110:19772–19777. doi: 10.1073/pnas.1314576110. - DOI - PMC - PubMed
-
- Barth M., Bryan R.K., Hegerl R. Approximation of missing-cone data in 3D electron microscopy. Ultramicroscopy. 1989;31:365–378. doi: 10.1016/0304-3991(89)90335-5. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
