

Modulation of virus-induced NF-κB signaling by NEMO coiled coil mimics
- PMID: 32286300
- PMCID: PMC7156456
- DOI: 10.1038/s41467-020-15576-3
Modulation of virus-induced NF-κB signaling by NEMO coiled coil mimics
Abstract
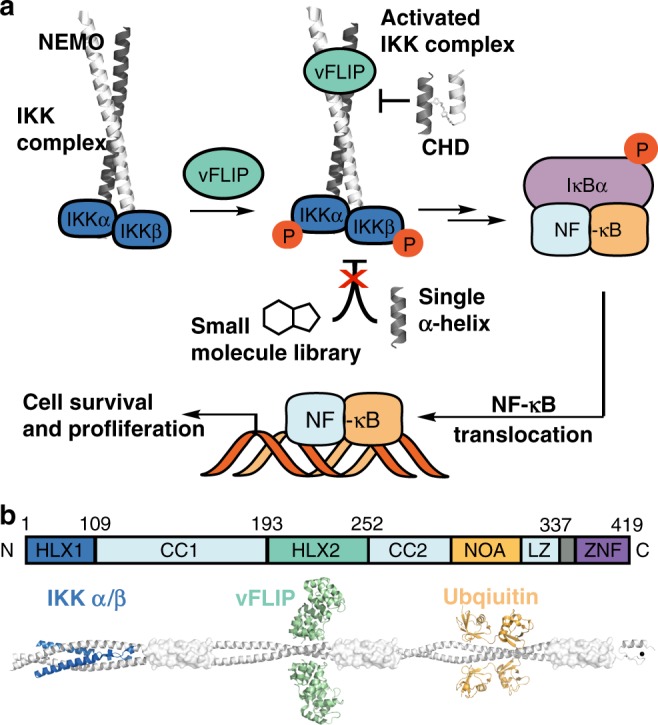
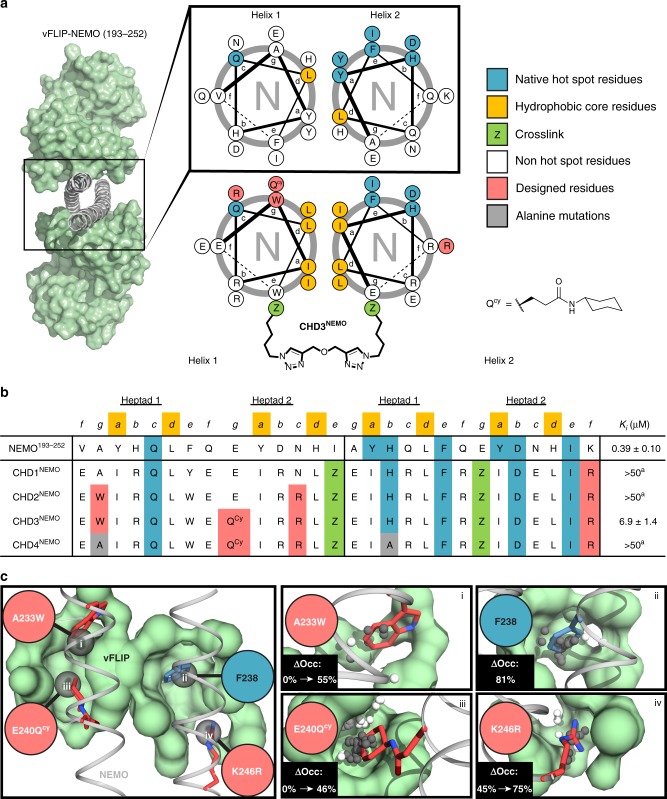
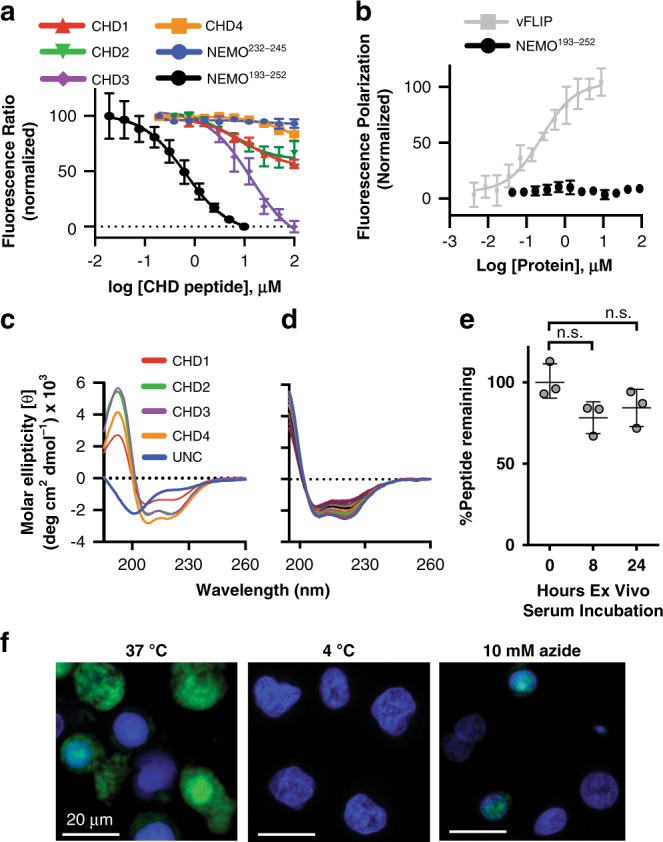
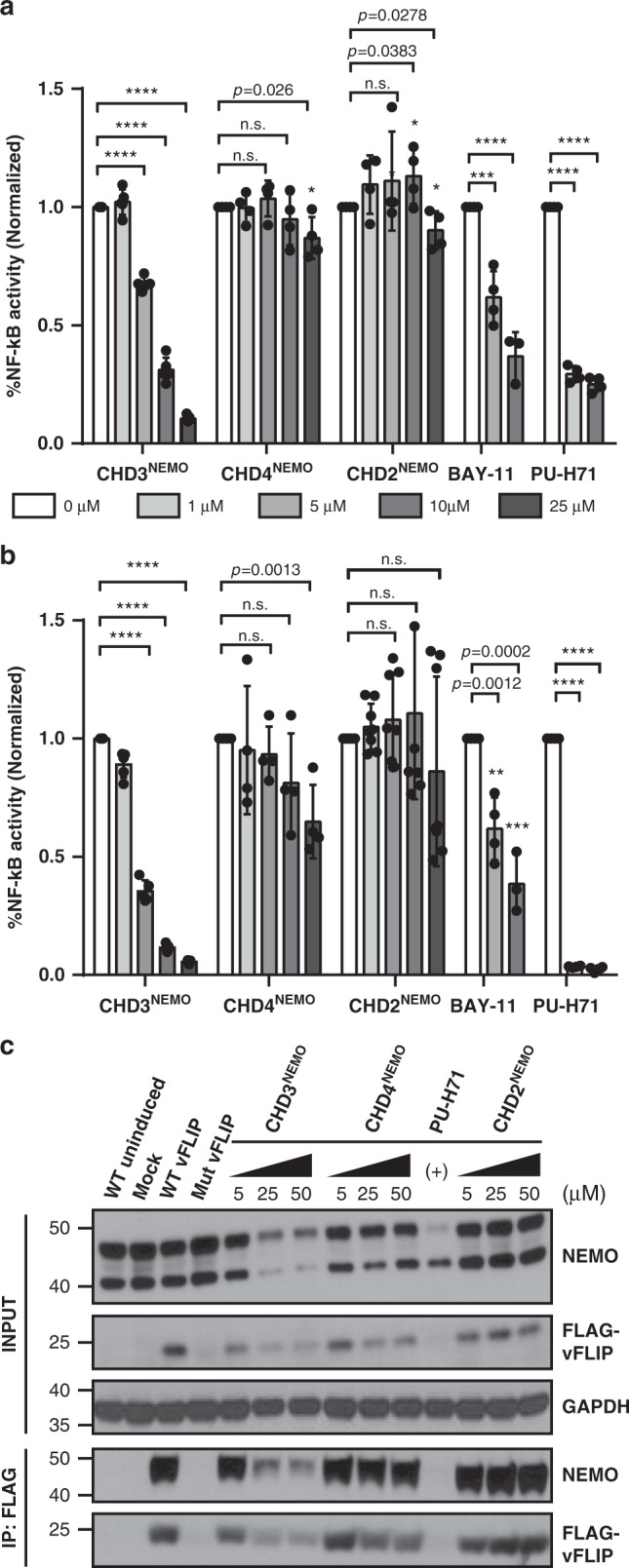
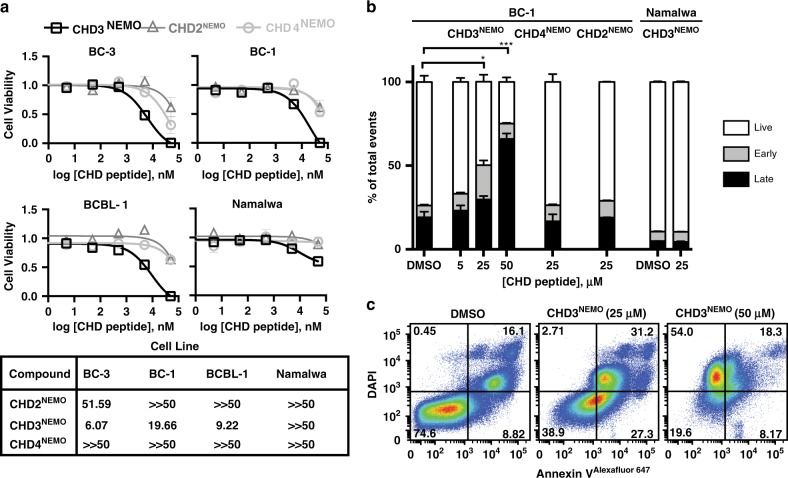
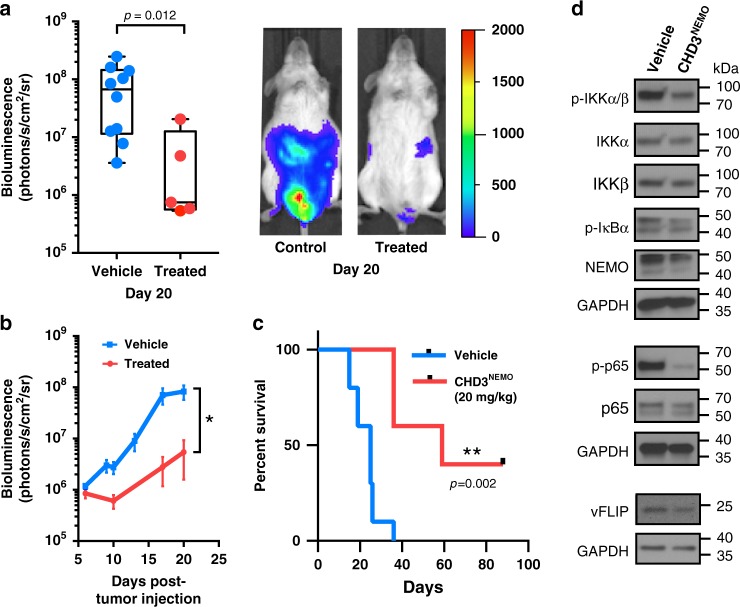
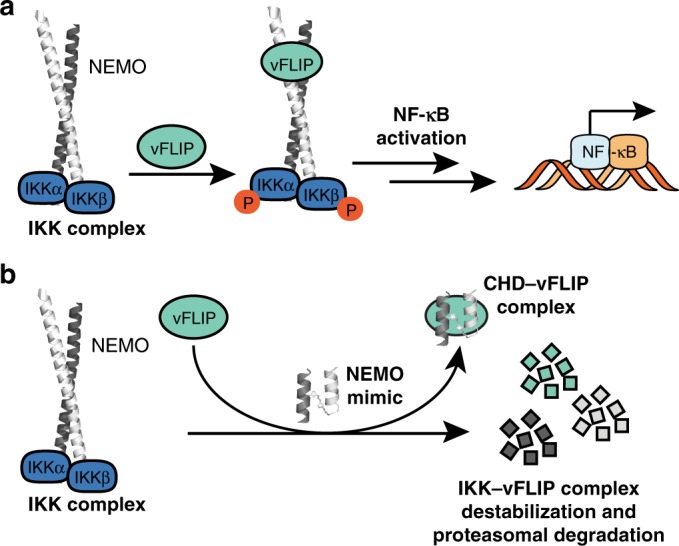
Protein-protein interactions featuring intricate binding epitopes remain challenging targets for synthetic inhibitors. Interactions of NEMO, a scaffolding protein central to NF-κB signaling, exemplify this challenge. Various regulators are known to interact with different coiled coil regions of NEMO, but the topological complexity of this protein has limited inhibitor design. We undertook a comprehensive effort to block the interaction between vFLIP, a Kaposi's sarcoma herpesviral oncoprotein, and NEMO using small molecule screening and rational design. Our efforts reveal that a tertiary protein structure mimic of NEMO is necessary for potent inhibition. The rationally designed mimic engages vFLIP directly causing complex disruption, protein degradation and suppression of NF-κB signaling in primary effusion lymphoma (PEL). NEMO mimic treatment induces cell death and delays tumor growth in a PEL xenograft model. Our studies with this inhibitor reveal the critical nexus of signaling complex stability in the regulation of NF-κB by a viral oncoprotein.
Conflict of interest statement
E.C., P.A., J.S. and M.W. are coinventors on a pending patent application (US62/768,373) that was filed jointly by Weill Cornell Medicine and New York University. The remaining authors declare no competing interests.
Figures
References
-
- Shimizu A, et al. Kaposi’s Sarcoma-Associated Herpesvirus vFLIP and Human T Cell Lymphotropic Virus Type 1 Tax Oncogenic Proteins Activate IκB Kinase Subunit γ by Different Mechanisms Independent of the Physiological Cytokine-Induced Pathways. J. Virol. 2011;85:7444–7448. doi: 10.1128/JVI.02337-10. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
