

Sex-specific innate immune selection of HIV-1 in utero is associated with increased female susceptibility to infection
- PMID: 32286302
- PMCID: PMC7156749
- DOI: 10.1038/s41467-020-15632-y
Sex-specific innate immune selection of HIV-1 in utero is associated with increased female susceptibility to infection
Erratum in
-
Author Correction: Sex-specific innate immune selection of HIV-1 in utero is associated with increased female susceptibility to infection.Nat Commun. 2020 May 4;11(1):2257. doi: 10.1038/s41467-020-16215-7. Nat Commun. 2020. PMID: 32367015 Free PMC article.
Abstract
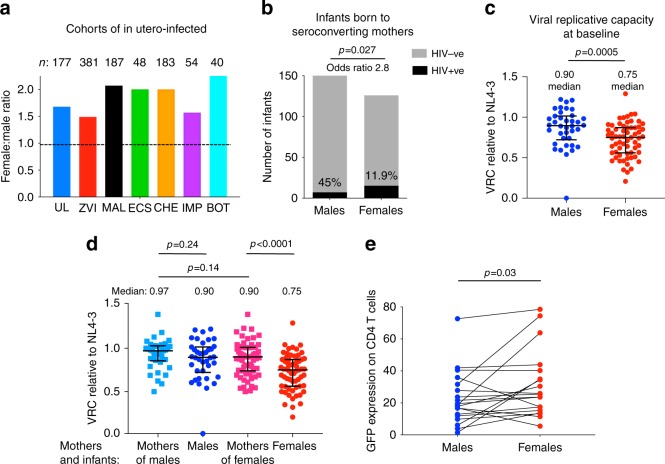
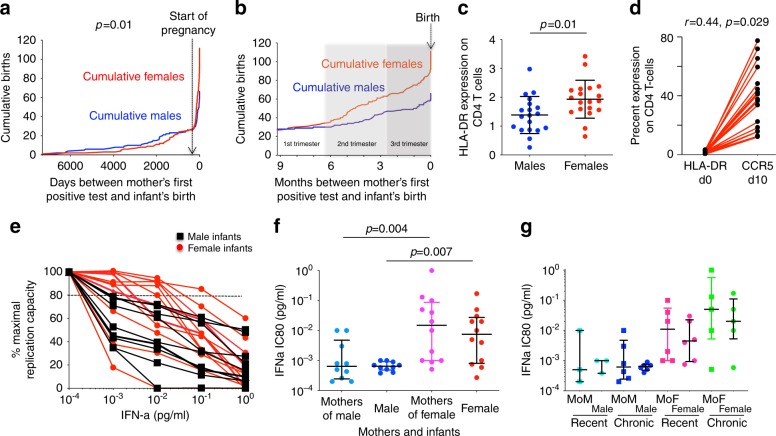
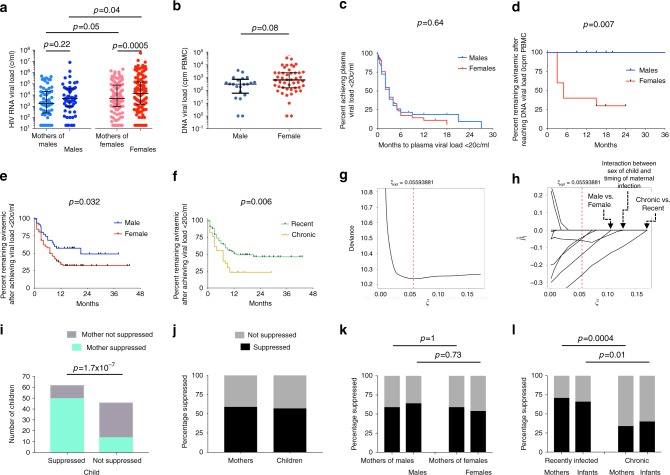
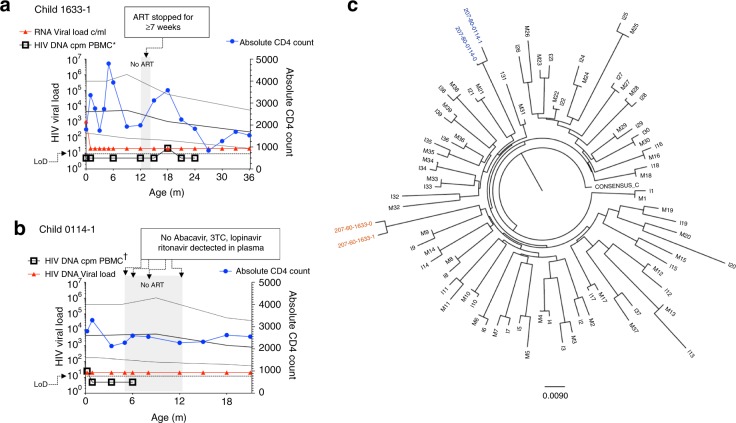
Female children and adults typically generate more efficacious immune responses to vaccines and infections than age-matched males, but also suffer greater immunopathology and autoimmune disease. We here describe, in a cohort of > 170 in utero HIV-infected infants from KwaZulu-Natal, South Africa, fetal immune sex differences resulting in a 1.5-2-fold increased female susceptibility to intrauterine HIV infection. Viruses transmitted to females have lower replicative capacity (p = 0.0005) and are more type I interferon-resistant (p = 0.007) than those transmitted to males. Cord blood cells from females of HIV-uninfected sex-discordant twins are more activated (p = 0.01) and more susceptible to HIV infection in vitro (p = 0.03). Sex differences in outcome include superior maintenance of aviraemia among males (p = 0.007) that is not explained by differential antiretroviral therapy adherence. These data demonstrate sex-specific innate immune selection of HIV associated with increased female susceptibility to in utero infection and enhanced functional cure potential among infected males.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Darwin, C. R. The Descent of Man and Selection in Relation to Sex (John Murray, London, 1871).
-
- Klein SL, Flanagan KL. Sex differences in immune responses. Nat. Rev. Immunol. 2016;16:626–638. - PubMed
-
- Flanagan KL, Fink AL, Plebanski M, Klein SL. Sex and gender differences in the outcomes of vaccination over the life course. Annu. Rev. Cell Dev. Biol. 2017;33:577–599. - PubMed
-
- Souyris M, et al. TLR7 escapes X chromosome inactivation in immune cells. Sci. Immunol. 2018;3:eaaqp8855. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
