

The Roles of Bacterial DNA Double-Strand Break Repair Proteins in Chromosomal DNA Replication
- PMID: 32286623
- PMCID: PMC7326373
- DOI: 10.1093/femsre/fuaa009
The Roles of Bacterial DNA Double-Strand Break Repair Proteins in Chromosomal DNA Replication
Abstract
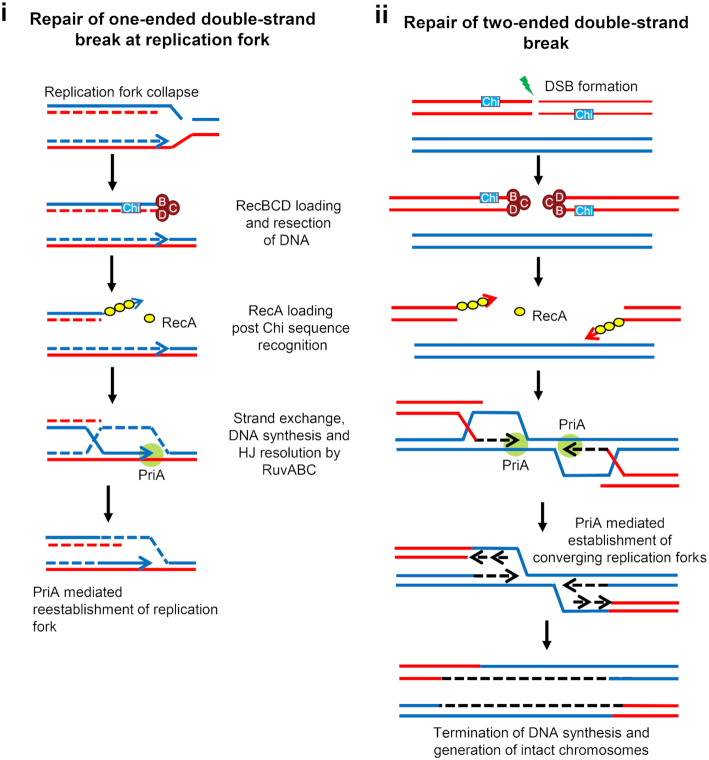
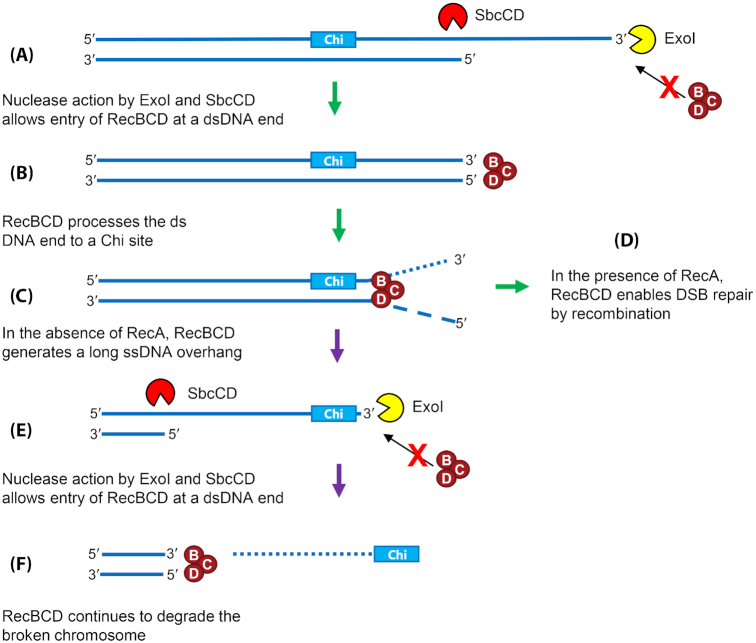
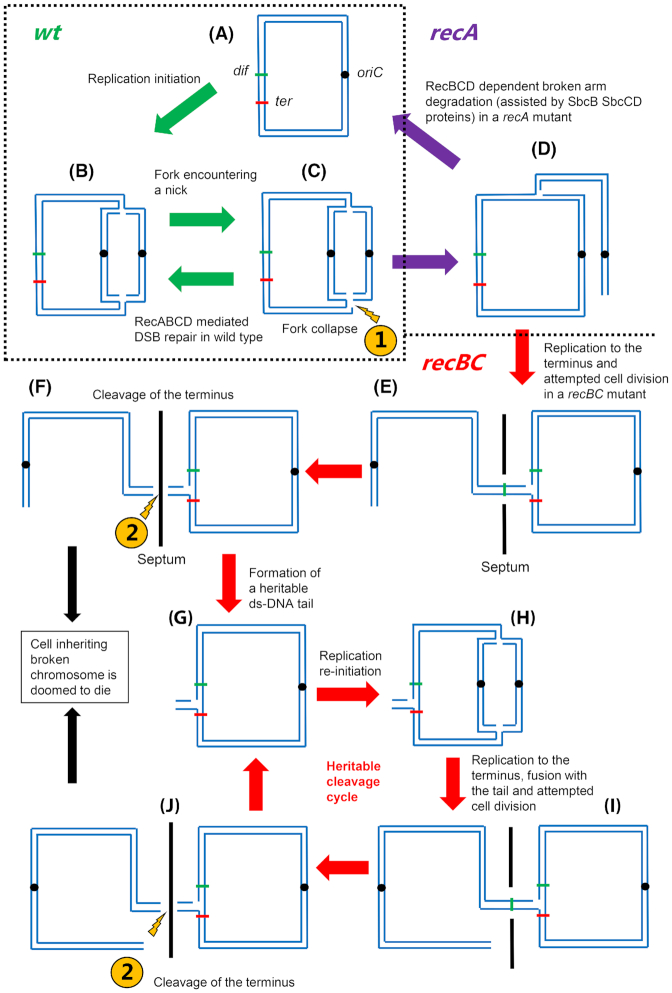
It is well established that DNA double-strand break (DSB) repair is required to underpin chromosomal DNA replication. Because DNA replication forks are prone to breakage, faithful DSB repair and correct replication fork restart are critically important. Cells, where the proteins required for DSB repair are absent or altered, display characteristic disturbances to genome replication. In this review, we analyze how bacterial DNA replication is perturbed in DSB repair mutant strains and explore the consequences of these perturbations for bacterial chromosome segregation and cell viability. Importantly, we look at how DNA replication and DSB repair processes are implicated in the striking recent observations of DNA amplification and DNA loss in the chromosome terminus of various mutant Escherichia coli strains. We also address the mutant conditions required for the remarkable ability to copy the entire E. coli genome, and to maintain cell viability, even in the absence of replication initiation from oriC, the unique origin of DNA replication in wild type cells. Furthermore, we discuss the models that have been proposed to explain these phenomena and assess how these models fit with the observed data, provide new insights and enhance our understanding of chromosomal replication and termination in bacteria.
Keywords: E. coli; DNA double-strand break repair; DNA replication; Homologous recombination.
© FEMS 2020.
Figures

References
-
- Anderson DG, Kowalczykowski SC. The translocating RecBCD enzyme stimulates recombination by directing RecA protein onto ssDNA in a chi-regulated manner. Cell. 1997;90:77–86. - PubMed
-
- Asai T, Imai M, Kogoma T. DNA damage-inducible replication of the Escherichia coli chromosome is initiated at separable sites within the minimal oriC. J Mol Biol. 1994;235:1459–69. - PubMed
