

S1P1 Contributes to Endotoxin-enhanced B-Cell Functions Involved in Hypersensitivity Pneumonitis
- PMID: 32289229
- PMCID: PMC7464516
- DOI: 10.1165/rcmb.2019-0339OC
S1P1 Contributes to Endotoxin-enhanced B-Cell Functions Involved in Hypersensitivity Pneumonitis
Abstract
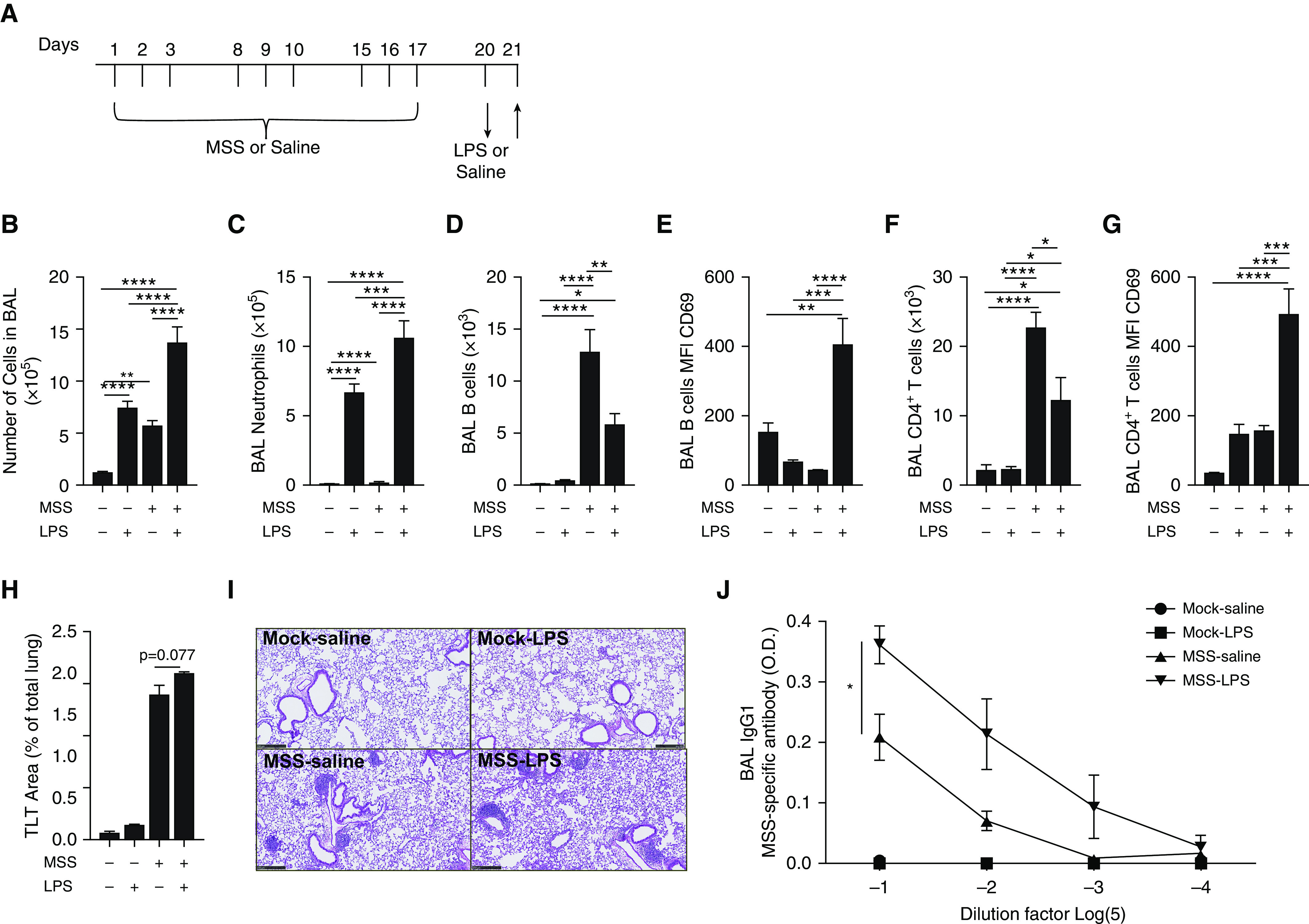
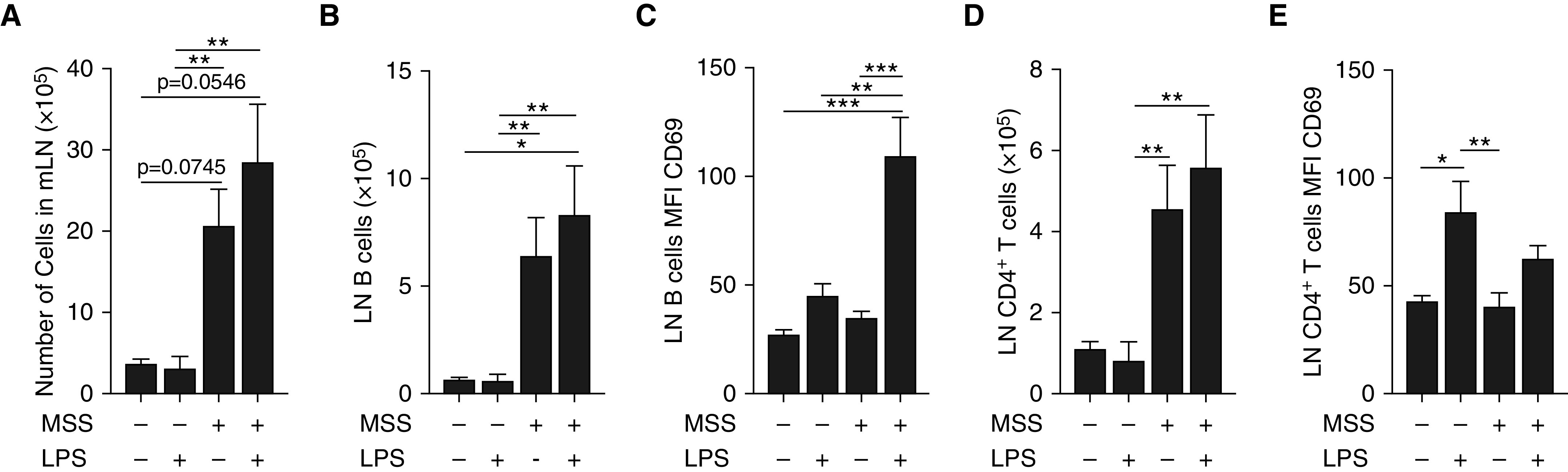
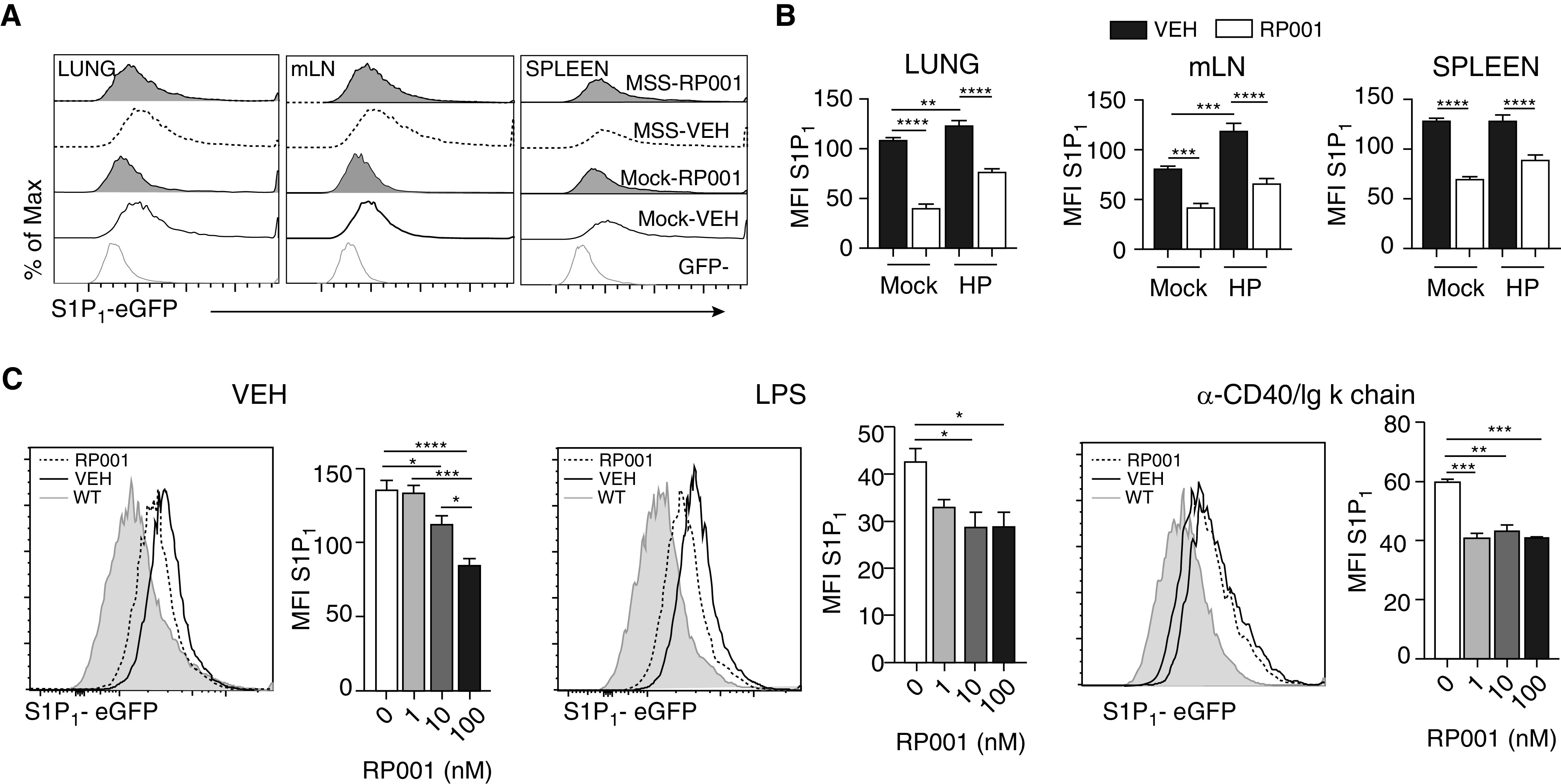
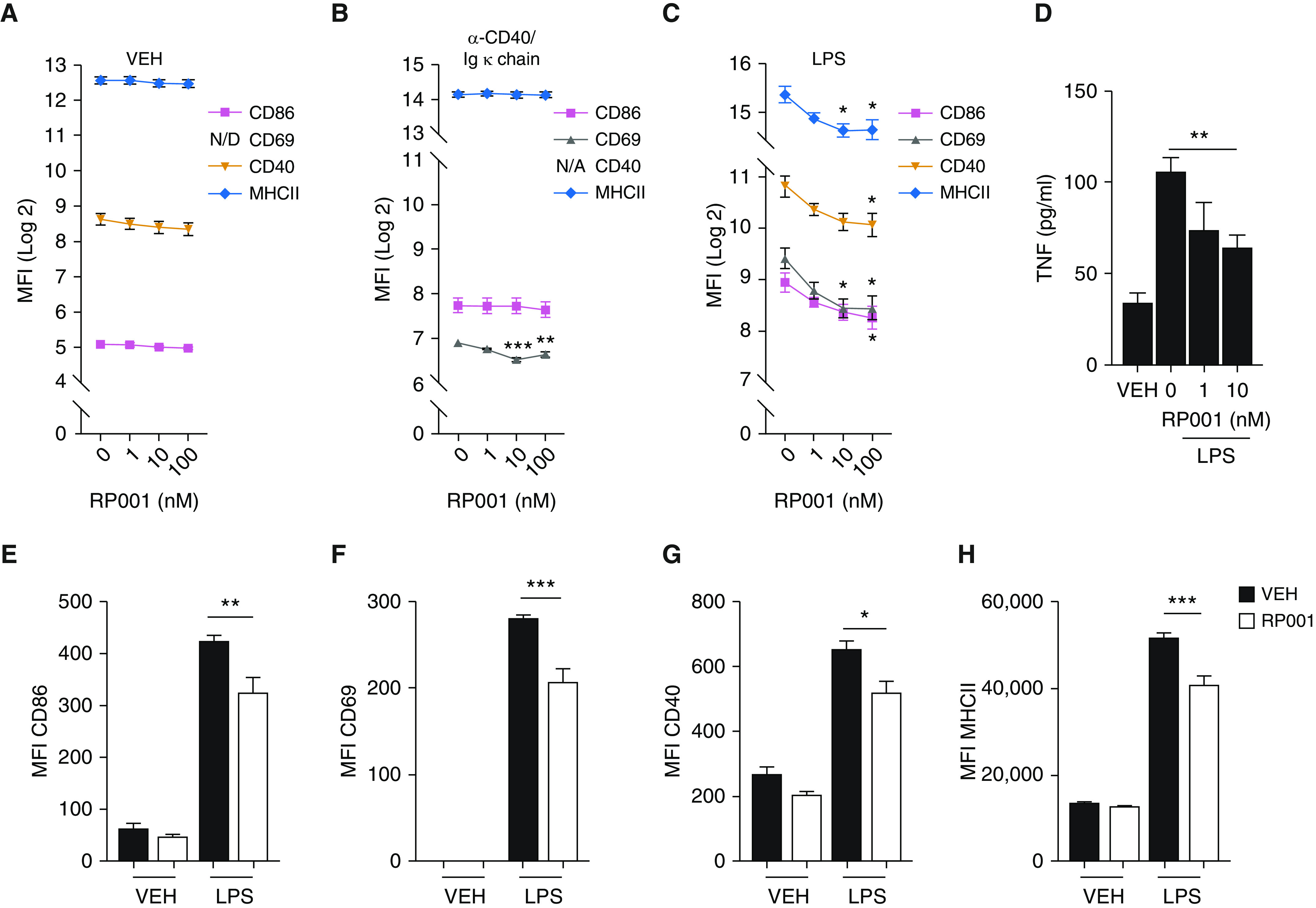
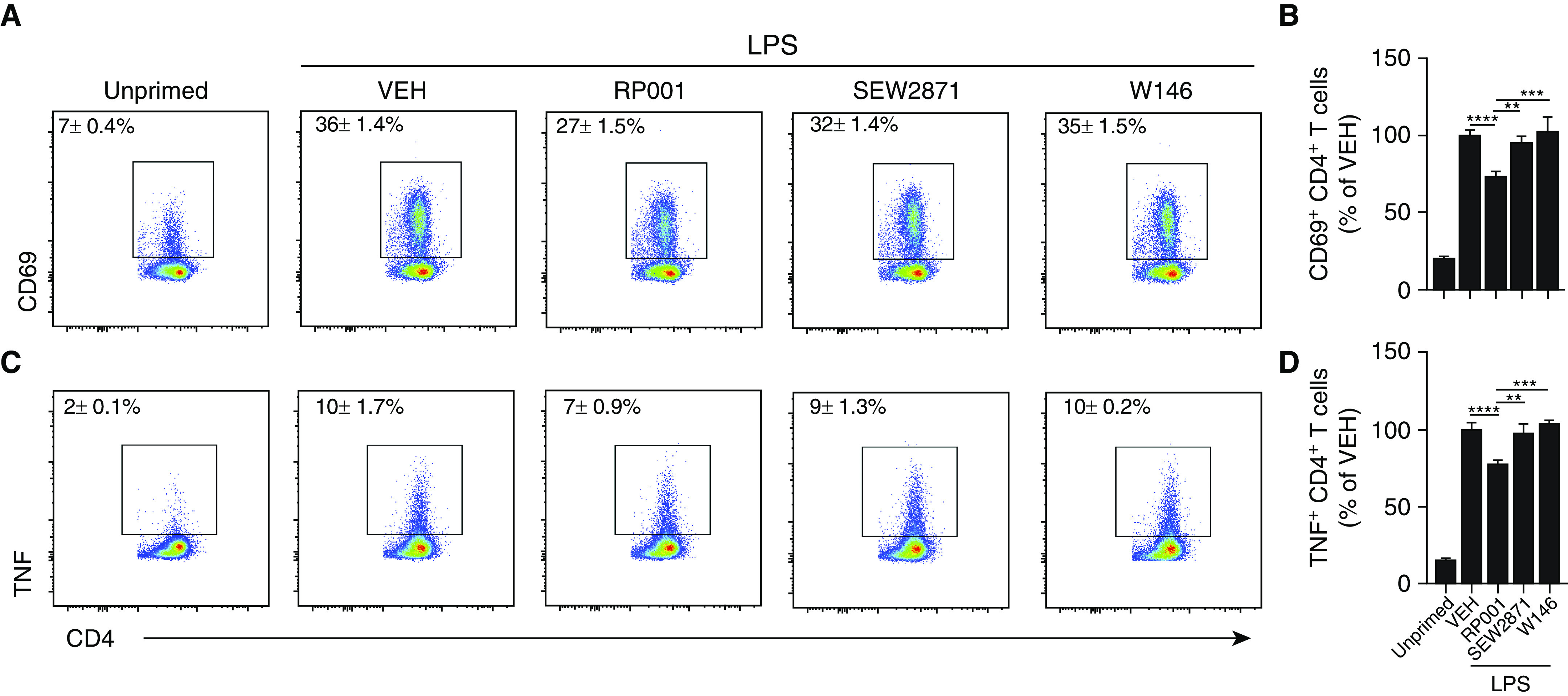
In a proportion of patients with hypersensitivity pneumonitis, the biological and environmental factors that sustain inflammation are ill defined, resulting in no effective treatment option. Bioaerosols found in occupational settings are complex and often include Toll-like receptor ligands, such as endotoxins. How Toll-like receptor ligands contribute to the persistence of hypersensitivity pneumonitis, however, remains poorly understood. In a previous study, we found that an S1P1 (sphingosine-1-phosphate receptor 1) agonist prevented the reactivation of antigen-driven B-cell responses in the lung. Here, we assessed the impact of endotoxins on B-cell activation in preexisting hypersensitivity pneumonitis and the role of S1P1 in this phenomenon. The impact of endotoxins on pre-established hypersensitivity pneumonitis was studied in vivo. S1P1 levels were tracked on B cells in the course of the disease using S1P1-eGFP knockin mice, and the role of S1P1 on B-cell functions was assessed using pharmacological tools. S1P1 was found on B cells in experimental hypersensitivity pneumonitis. Endotoxin exposure enhanced neutrophil accumulation in the BAL of mice with experimental hypersensitivity pneumonitis. This was associated with enhanced CD69 cell-surface expression on lymphocytes in the BAL. In isolated B cells, endotoxins increased cell-surface levels of costimulatory molecules and CD69, which was prevented by an S1P1 agonist. S1P1 modulators also reduced TNF production by B cells and their capacity to trigger T-cell cooperation ex vivo. An S1P1 ligand directly inhibited endotoxin-induced B-cell activation.
Keywords: TLR4; TNF; antibodies; extrinsic allergic alveolitis; ozanimod.
Figures

References
-
- Lacasse Y, Girard M, Cormier Y. Recent advances in hypersensitivity pneumonitis. Chest. 2012;142:208–217. - PubMed
-
- Suda T, Chida K, Hayakawa H, Imokawa S, Iwata M, Nakamura H, et al. Development of bronchus-associated lymphoid tissue in chronic hypersensitivity pneumonitis. Chest. 1999;115:357–363. - PubMed
-
- Hanak V, Golbin JM, Ryu JH. Causes and presenting features in 85 consecutive patients with hypersensitivity pneumonitis. Mayo Clin Proc. 2007;82:812–816. - PubMed
-
- Fernández Pérez ER, Kong AM, Raimundo K, Koelsch TL, Kulkarni R, Cole AL. Epidemiology of hypersensitivity pneumonitis among an insured population in the United States: a claims-based cohort analysis. Ann Am Thorac Soc. 2018;15:460–469. - PubMed
-
- Walters GI, Mokhlis JM, Moore VC, Robertson AS, Burge GA, Bhomra PS, et al. Characteristics of hypersensitivity pneumonitis diagnosed by interstitial and occupational lung disease multi-disciplinary team consensus. Respir Med. 2019;155:19–25. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
