

A Comprehensive Transcriptome Analysis Identifies FXN and BDNF as Novel Targets of miRNAs in Friedreich's Ataxia Patients
- PMID: 32291635
- PMCID: PMC7253519
- DOI: 10.1007/s12035-020-01899-1
A Comprehensive Transcriptome Analysis Identifies FXN and BDNF as Novel Targets of miRNAs in Friedreich's Ataxia Patients
Abstract
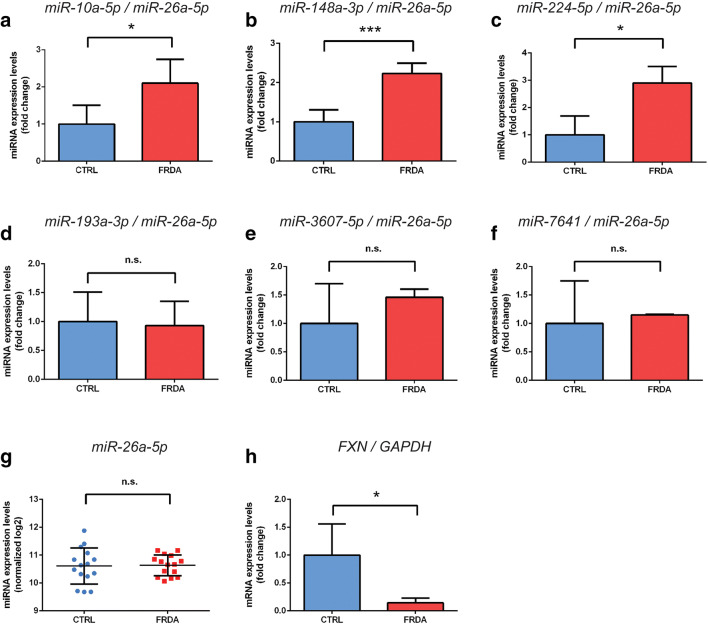
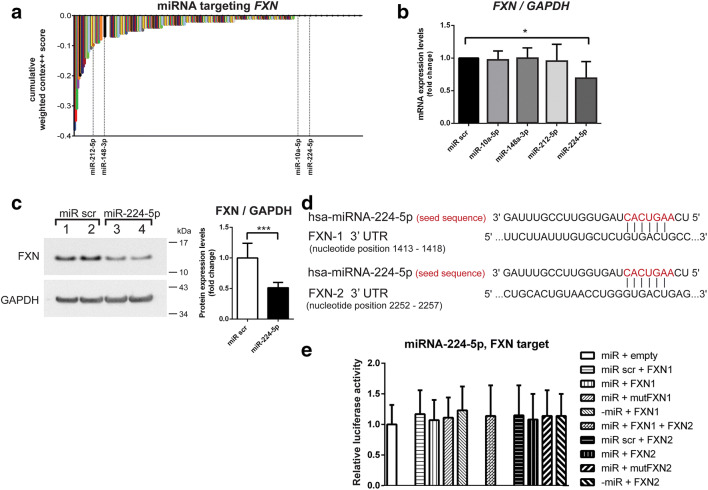
Friedreich's ataxia (FRDA) is a genetic neurodegenerative disease that is caused by guanine-adenine-adenine (GAA) nucleotide repeat expansions in the first intron of the frataxin (FXN) gene. Although present in the intron, this mutation leads to a substantial decrease in protein expression. Currently, no effective treatment is available for FRDA, and, in addition to FXN, other targets with therapeutic potential are continuously sought. As miRNAs can regulate the expression of a broad spectrum of genes, are used as biomarkers, and can serve as therapeutic tools, we decided to identify and characterize differentially expressed miRNAs and their targets in FRDA cells compared to unaffected control (CTRL) cells. In this study, we performed an integrated miRNAseq and RNAseq analysis using the same cohort of primary FRDA and CTRL cells. The results of the transcriptome studies were supported by bioinformatic analyses and validated by qRT-PCR. miRNA interactions with target genes were assessed by luciferase assays, qRT-PCR, and immunoblotting. In silico analysis identified the FXN transcript as a target of five miRNAs upregulated in FRDA cells. Further studies confirmed that miRNA-224-5p indeed targets FXN, resulting in decreases in mRNA and protein levels. We also validated the ability of miRNA-10a-5p to bind and regulate the levels of brain-derived neurotrophic factor (BDNF), an important modulator of neuronal growth. We observed a significant decrease in the levels of miRNA-10a-5p and increase in the levels of BDNF upon correction of FRDA cells via zinc-finger nuclease (ZFN)-mediated excision of expanded GAA repeats. Our comprehensive transcriptome analyses identified miRNA-224-5p and miRNA-10a-5p as negative regulators of the FXN and BDNF expression, respectively. These results emphasize not only the importance of miRNAs in the pathogenesis of FRDA but also their potential as therapeutic targets for this disease.
Keywords: Brain-derived neurotrophic factor (BDNF); Frataxin (FXN); Friedreich’s ataxia (FRDA); RNAseq; miRNAseq; microRNA-10a-5p; microRNA-224-5p.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures




References
-
- Marmolino D. Friedreich’s ataxia: past, present and future. Brain Res Rev. 2011;67(1–2):311–330. - PubMed
-
- Delatycki MB, Bidichandani SI. Friedreich ataxia- pathogenesis and implications for therapies. Neurobiol Dis. 2019;132:104606. - PubMed
-
- Li Y, Lu Y, Polak U, Lin K, Shen J, Farmer J, Seyer L, Bhalla AD, Rozwadowska N, Lynch DR, Butler JS, Napierala M. Expanded GAA repeats impede transcription elongation through the FXN gene and induce transcriptional silencing that is restricted to the FXN locus. Hum Mol Genet. 2015;24(24):6932–6943. - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
