

The Design and Development of a Multi-HBV Antigen Encoded in Chimpanzee Adenoviral and Modified Vaccinia Ankara Viral Vectors; A Novel Therapeutic Vaccine Strategy against HBV
- PMID: 32295168
- PMCID: PMC7348829
- DOI: 10.3390/vaccines8020184
The Design and Development of a Multi-HBV Antigen Encoded in Chimpanzee Adenoviral and Modified Vaccinia Ankara Viral Vectors; A Novel Therapeutic Vaccine Strategy against HBV
Abstract
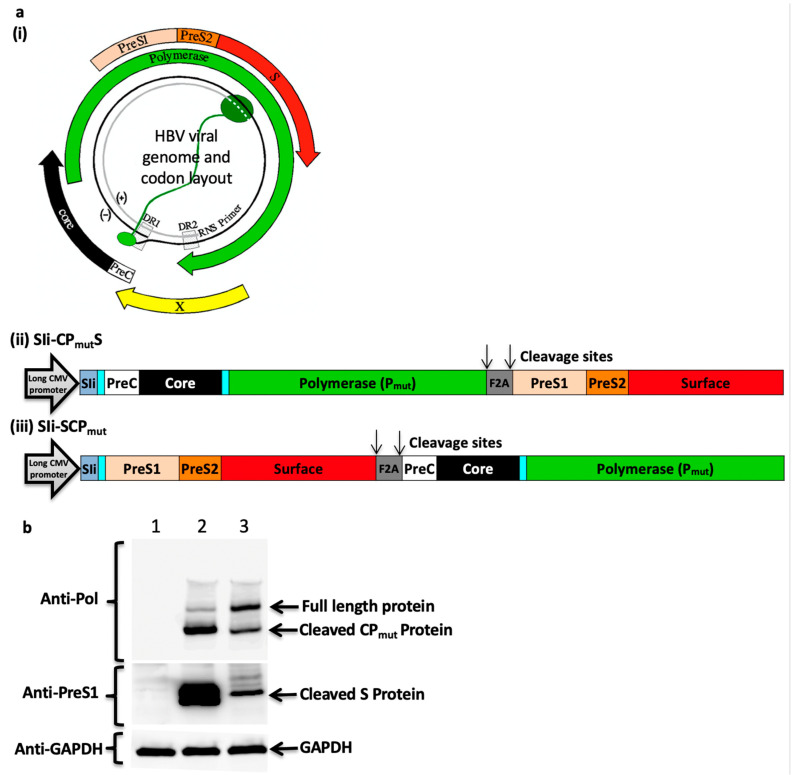
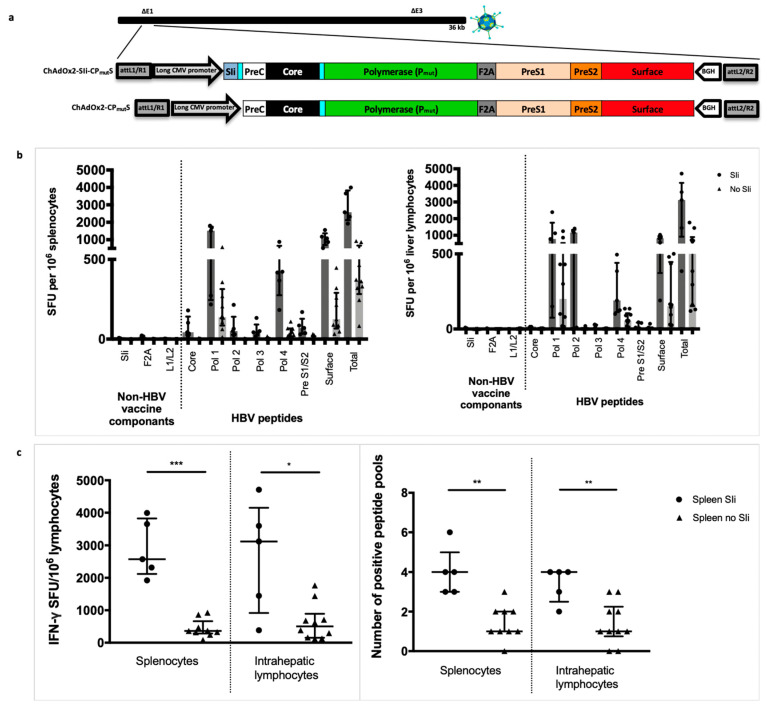
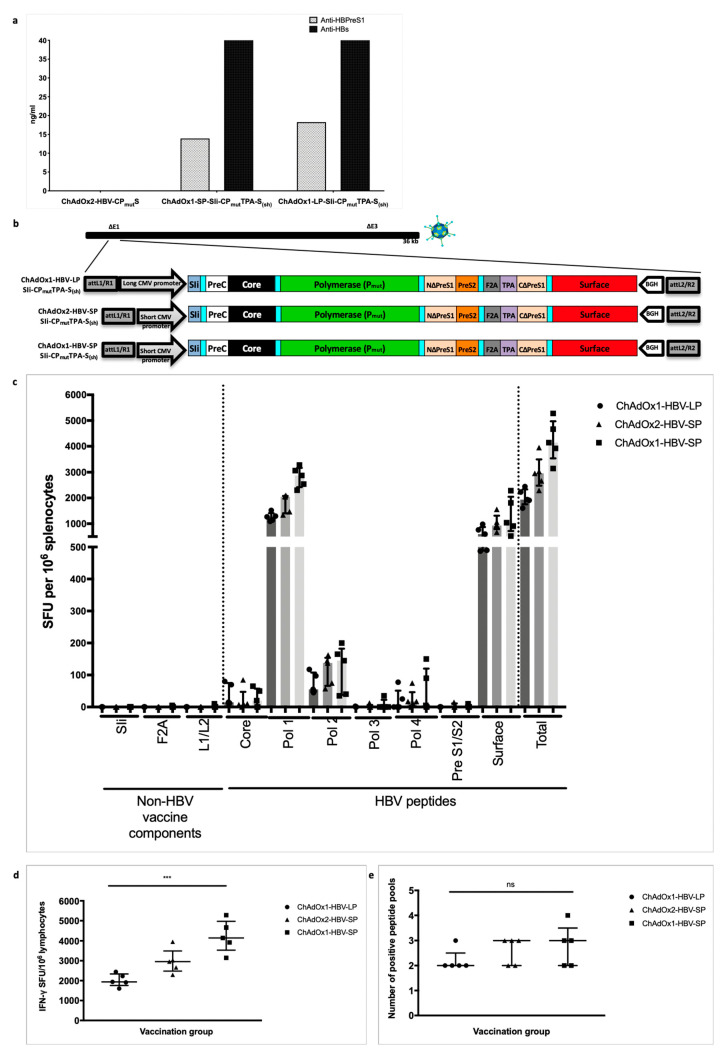
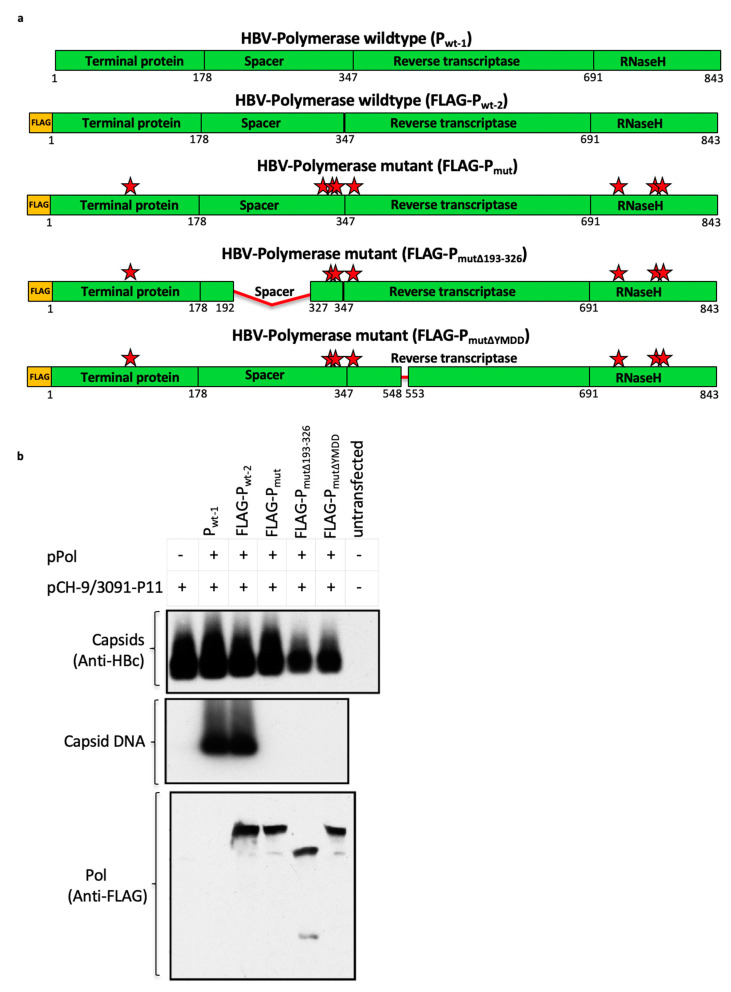
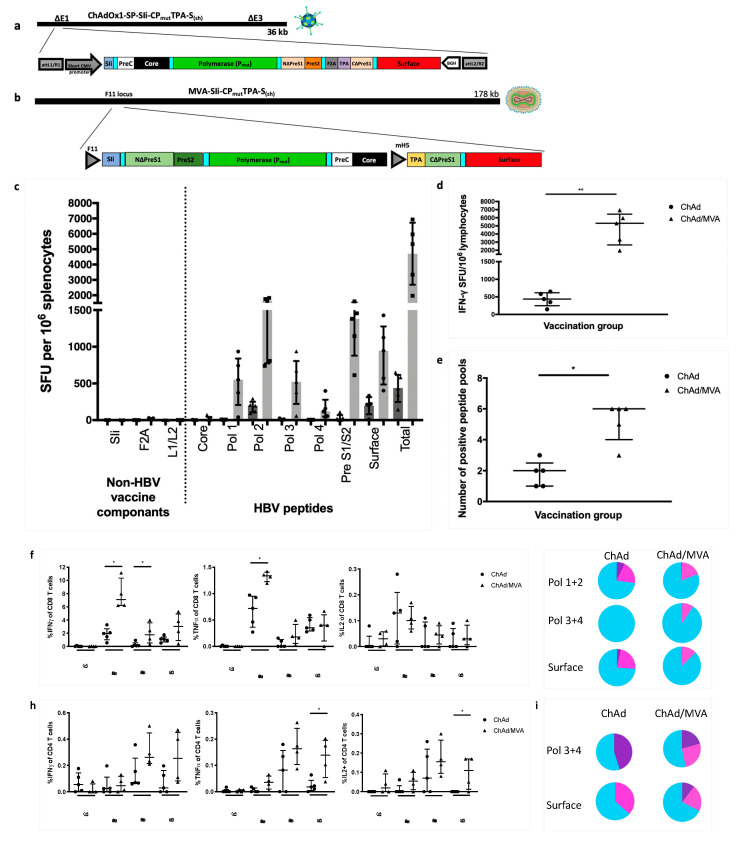
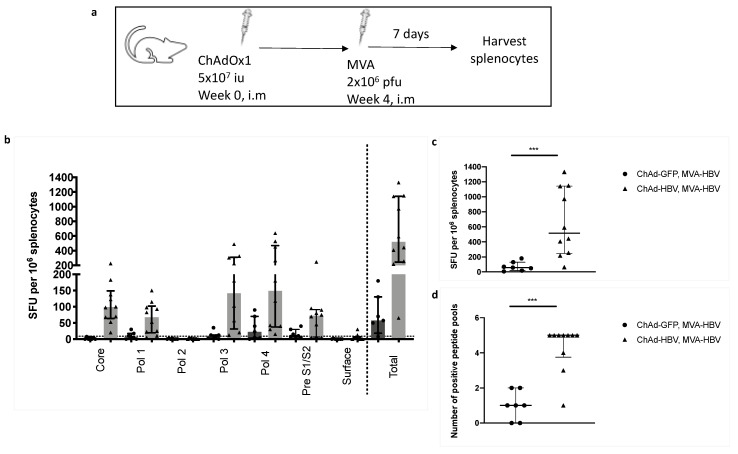
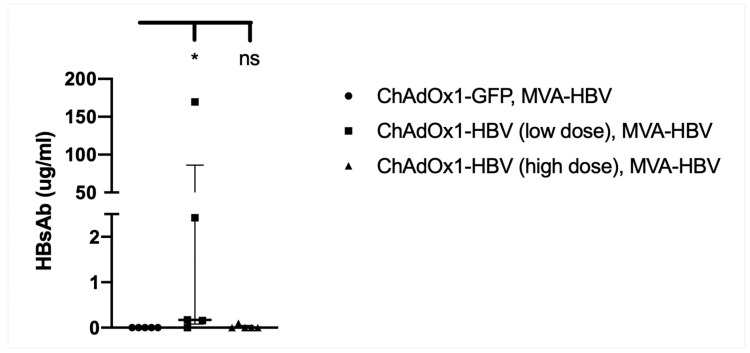
Chronic hepatitis B virus (HBV) infection affects 257 million people globally. Current therapies suppress HBV but viral rebound occurs on cessation of therapy; novel therapeutic strategies are urgently required. To develop a therapeutic HBV vaccine that can induce high magnitude T cells to all major HBV antigens, we have developed a novel HBV vaccine using chimpanzee adenovirus (ChAd) and modified vaccinia Ankara (MVA) viral vectors encoding multiple HBV antigens. ChAd vaccine alone generated very high magnitude HBV specific T cell responses to all HBV major antigens. The inclusion of a shark Invariant (SIi) chain genetic adjuvant significantly enhanced the magnitude of T-cells against HBV antigens. Compared to ChAd alone vaccination, ChAd-prime followed by MVA-boost vaccination further enhanced the magnitude and breadth of the vaccine induced T cell response. Intra-cellular cytokine staining study showed that HBV specific CD8+ and CD4+ T cells were polyfunctional, producing combinations of IFNγ, TNF-α, and IL-2. In summary, we have generated genetically adjuvanted ChAd and MVA vectored HBV vaccines with the potential to induce high-magnitude T cell responses through a prime-boost therapeutic vaccination approach. These pre-clinical studies pave the way for new studies of HBV therapeutic vaccination in humans with chronic hepatitis B infection.
Keywords: ChAd; ChAdOx1; Hepatitis B virus (HBV); MVA; T cell vaccine; chimpanzee adenovirus; modified vaccinia Ankara; therapeutic HBV vaccine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Chan H.L., Chan C.K., Hui A.J., Chan S., Poordad F., Chang T.T., Mathurin P., Flaherty J.F., Lin L., Corsa A., et al. Effects of tenofovir disoproxil fumarate in hepatitis B e antigen-positive patients with normal levels of alanine aminotransferase and high levels of hepatitis B virus DNA. Gastroenterology. 2014;146:1240–1248. doi: 10.1053/j.gastro.2014.01.044. - DOI - PubMed
-
- Li M.R., Xi H.L., Wang Q.H., Hou F.Q., Huo N., Zhang X.X., Li F., Xu X.Y. Kinetics and prediction of HBsAg loss during long-term therapy with nucleos(t)ide analogues of different potency in patients with chronic hepatitis B. PLoS ONE. 2014;9:e98476. doi: 10.1371/journal.pone.0098476. - DOI - PMC - PubMed
-
- Wu C.Y., Lin J.T., Ho H.J., Su C.W., Lee T.Y., Wang S.Y., Wu C., Wu J.C. Association of nucleos(t)ide analogue therapy with reduced risk of hepatocellular carcinoma in patients with chronic hepatitis B: A nationwide cohort study. Gastroenterology. 2014;147:143–151.e145. doi: 10.1053/j.gastro.2014.03.048. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
