

Silencing MYH9 blocks HBx-induced GSK3β ubiquitination and degradation to inhibit tumor stemness in hepatocellular carcinoma
- PMID: 32296025
- PMCID: PMC7018736
- DOI: 10.1038/s41392-020-0111-4
Silencing MYH9 blocks HBx-induced GSK3β ubiquitination and degradation to inhibit tumor stemness in hepatocellular carcinoma
Abstract
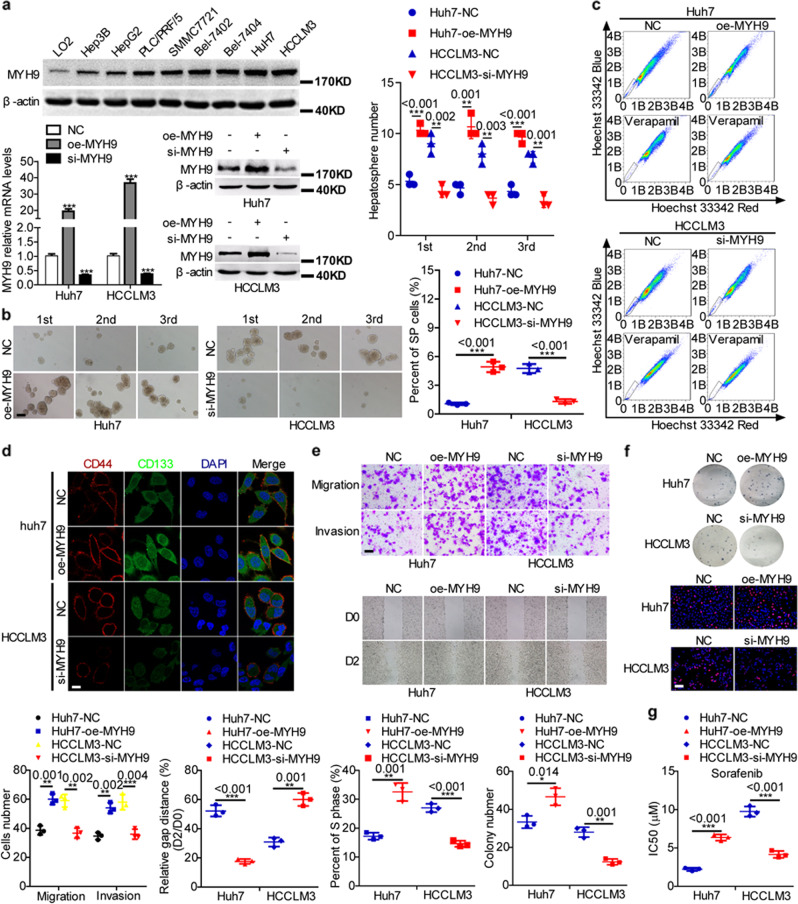
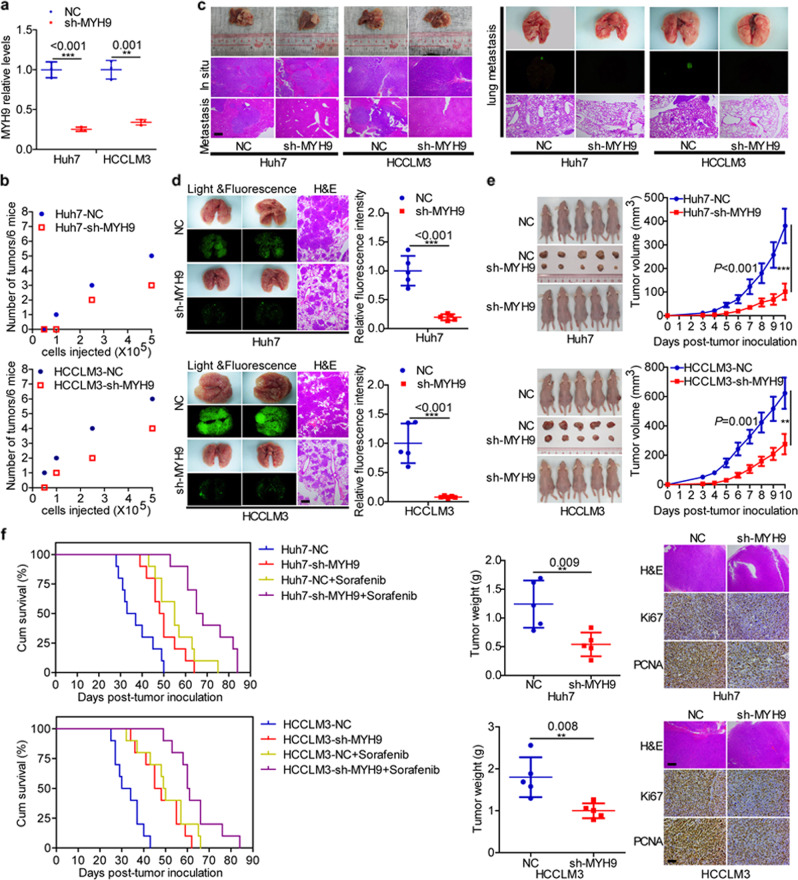
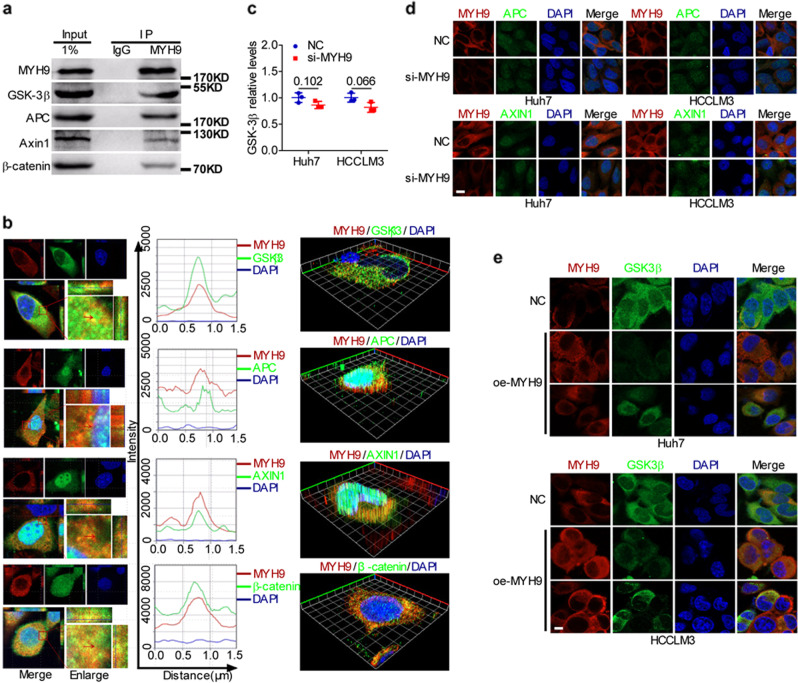
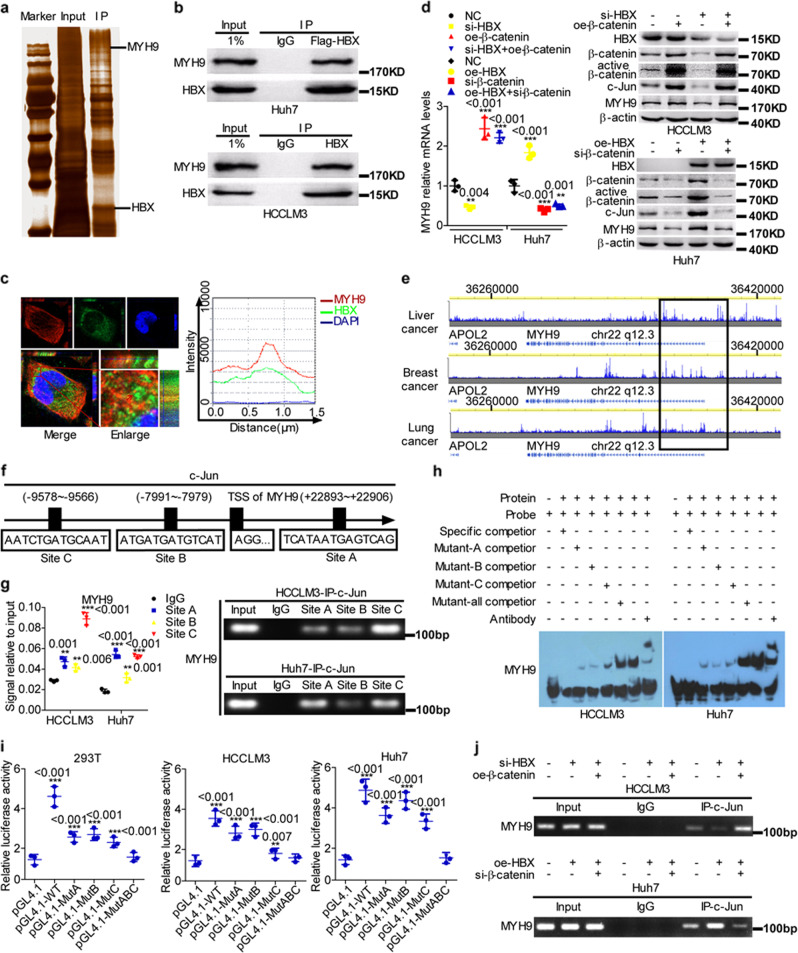
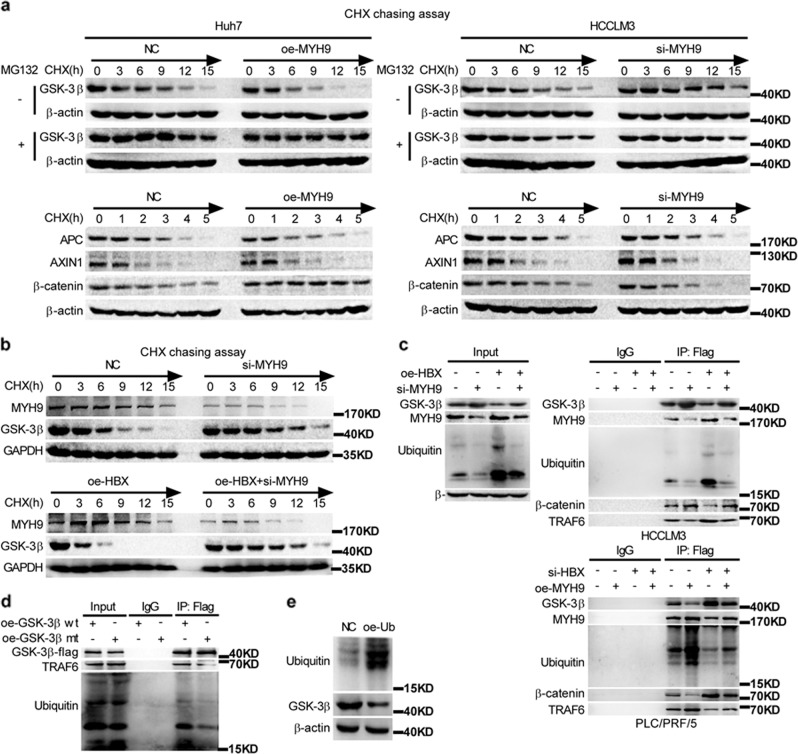
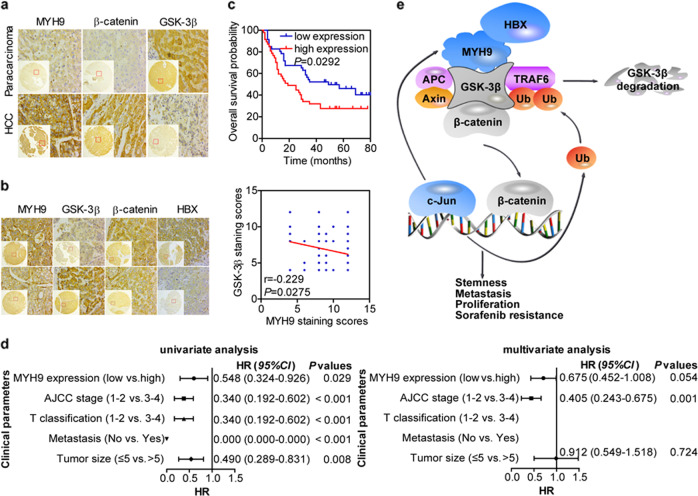
MYH9 has dual functions in tumors. However, its role in inducing tumor stemness in hepatocellular carcinoma (HCC) is not yet determined. Here, we found that MYH9 is an effective promoter of tumor stemness that facilitates hepatocellular carcinoma pathogenesis. Importantly, targeting MYH9 remarkably improved the survival of hepatocellular carcinoma-bearing mice and promoted sorafenib sensitivity of hepatocellular carcinoma cells in vivo. Mechanistic analysis suggested that MYH9 interacted with GSK3β and reduced its protein expression by ubiquitin-mediated degradation, which therefore dysregulated the β-catenin destruction complex and induced the downstream tumor stemness phenotype, epithelial-mesenchymal transition, and c-Jun signaling in HCC. C-Jun transcriptionally stimulated MYH9 expression and formed an MYH9/GSK3β/β-catenin/c-Jun feedback loop. X protein is a hepatitis B virus (HBV)-encoded key oncogenic protein that promotes HCC pathogenesis. Interestingly, we observed that HBV X protein (HBX) interacted with MYH9 and induced its expression by modulating GSK3β/β-catenin/c-Jun signaling. Targeting MYH9 blocked HBX-induced GSK3β ubiquitination to activate the β-catenin destruction complex and suppressed cancer stemness and EMT. Based on TCGA database analysis, MYH9 was found to be elevated and conferred poor prognosis for hepatocellular carcinoma patients. In clinical samples, high MYH9 expression levels predicted poor prognosis of hepatocellular carcinoma patients. These findings identify the suppression of MYH9 as an alternative approach for the effective eradication of CSC properties to inhibit cancer migration, invasion, growth, and sorafenib resistance in HCC patients. Our study demonstrated that MYH9 is a crucial therapeutic target in HCC.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
