

Germline Elongator mutations in Sonic Hedgehog medulloblastoma
- PMID: 32296180
- PMCID: PMC7430762
- DOI: 10.1038/s41586-020-2164-5
Germline Elongator mutations in Sonic Hedgehog medulloblastoma
Abstract
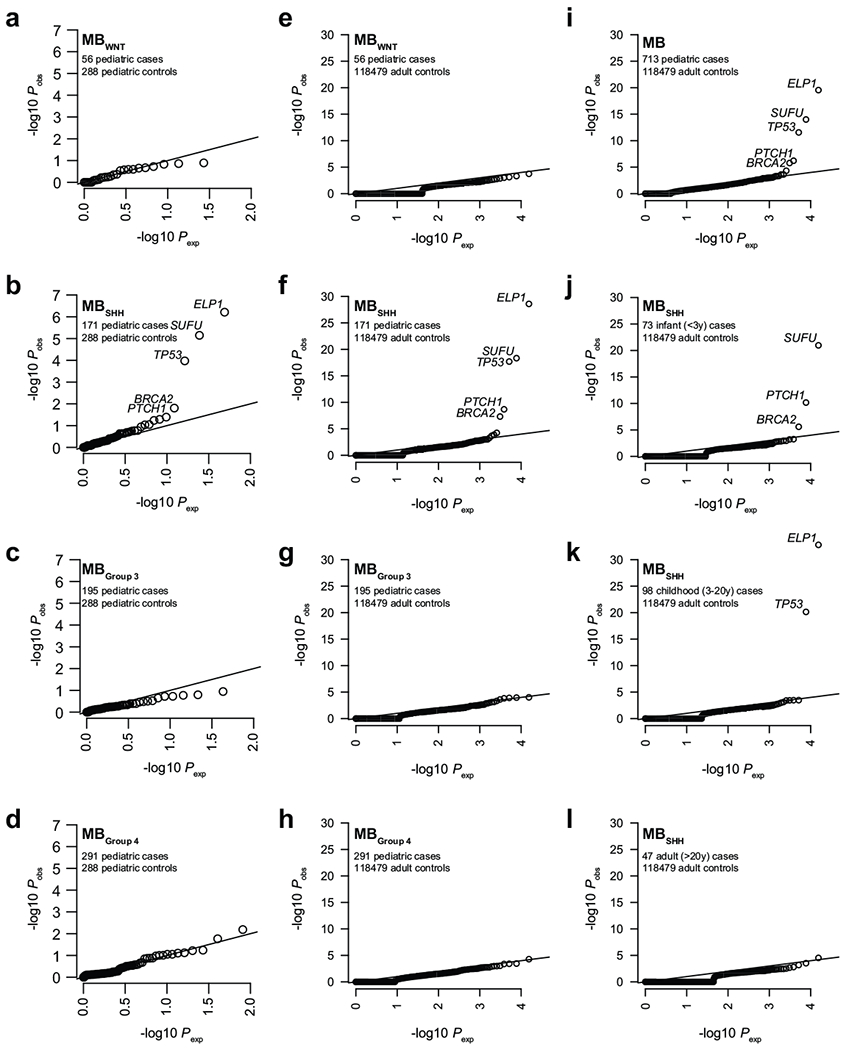
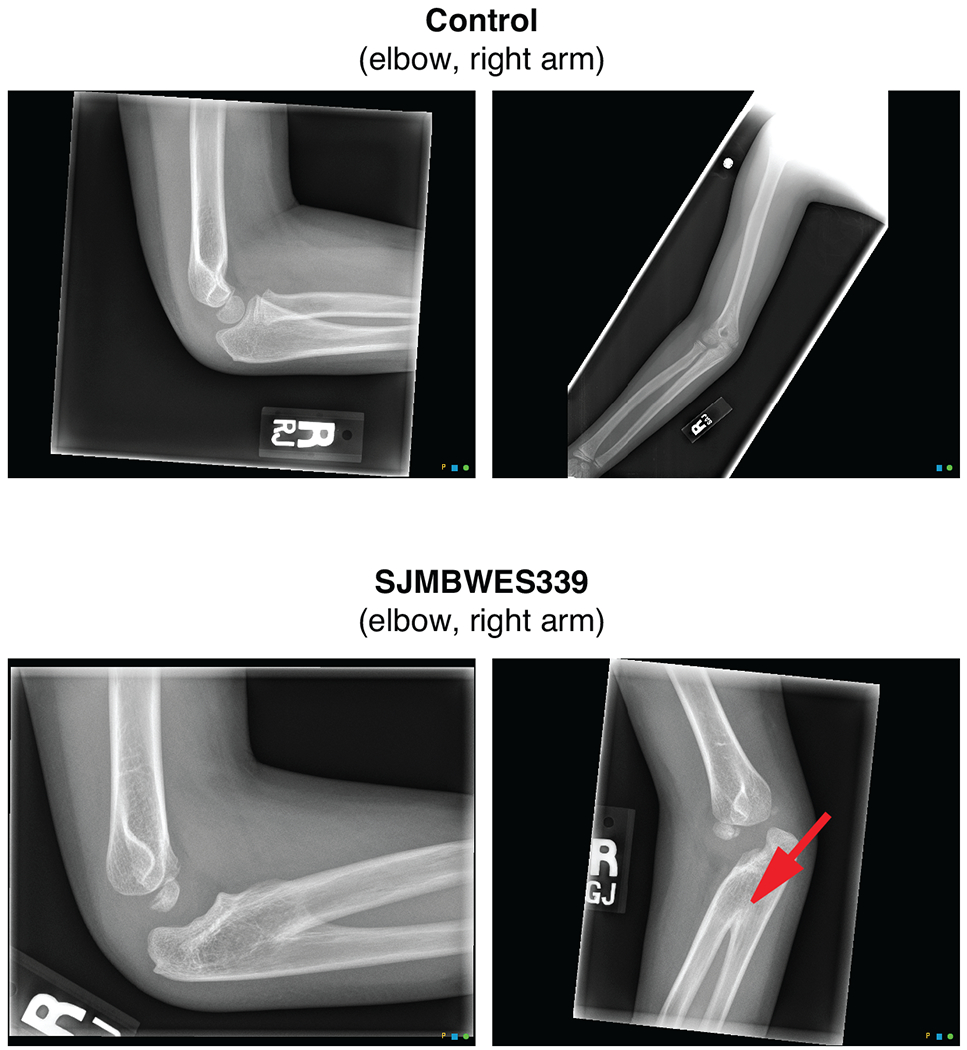
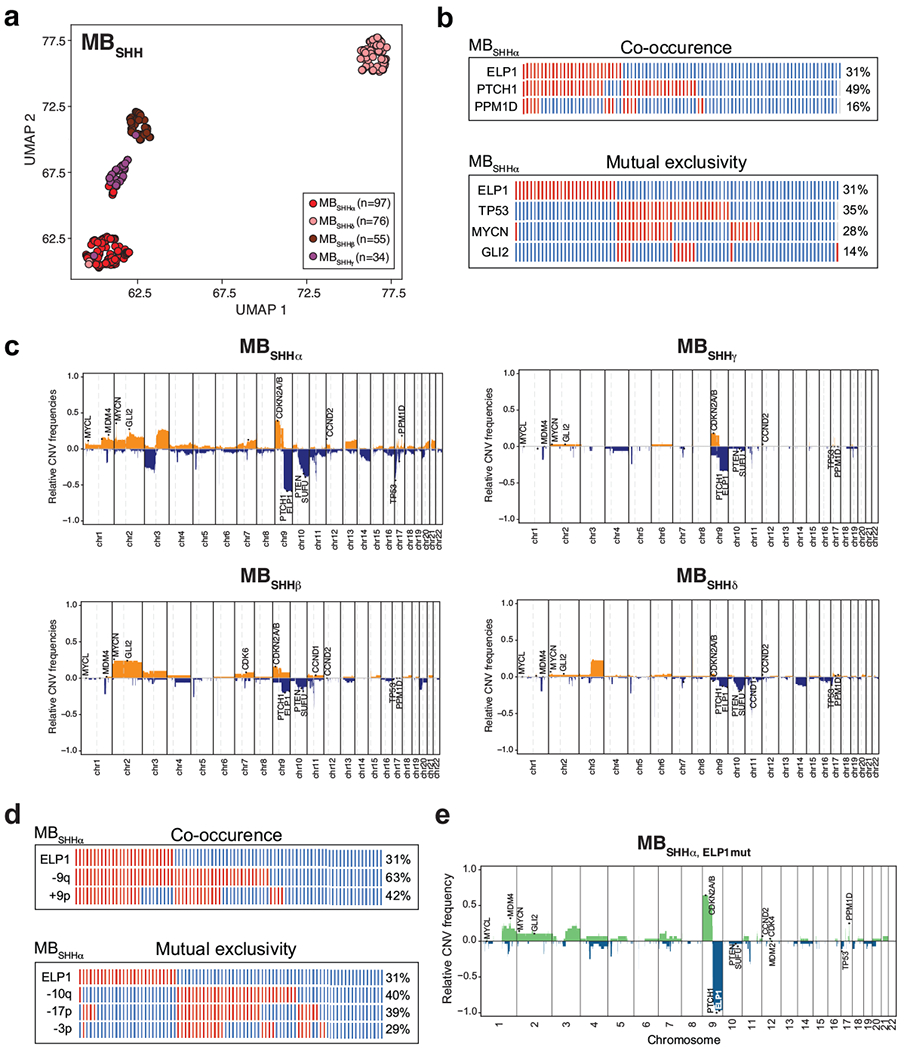
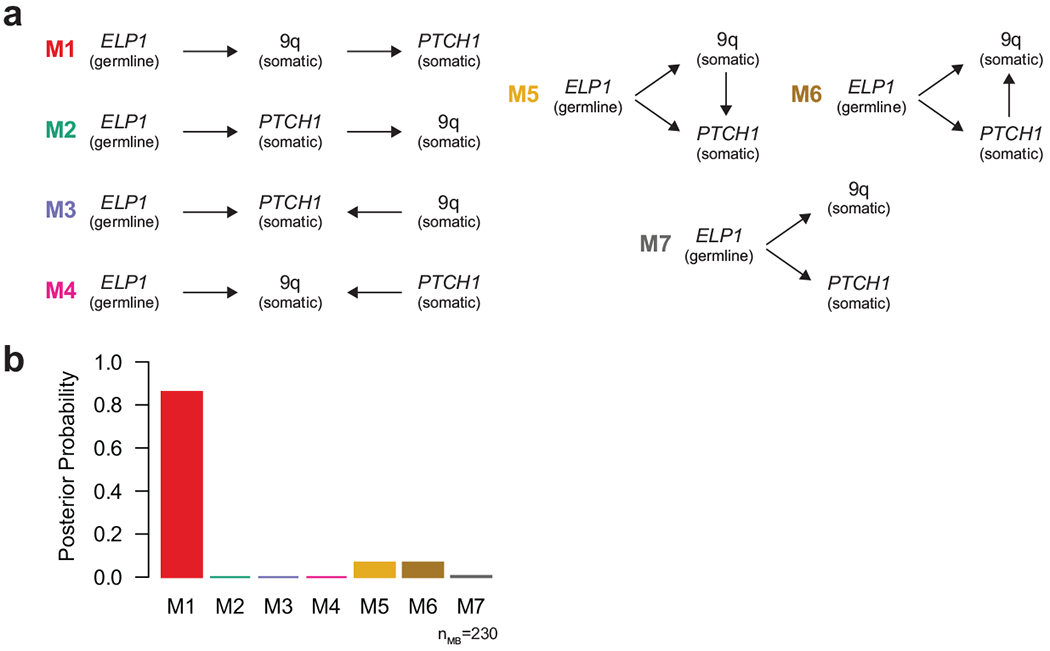
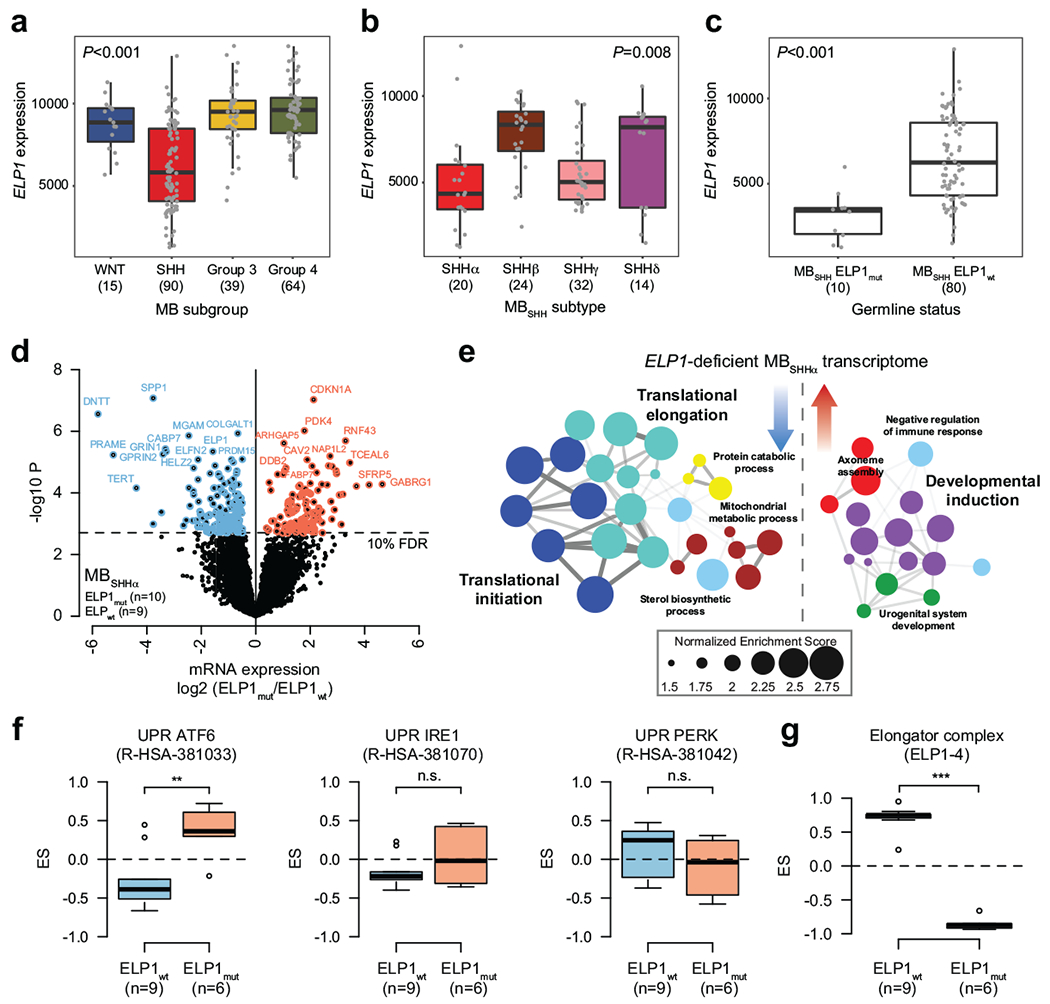
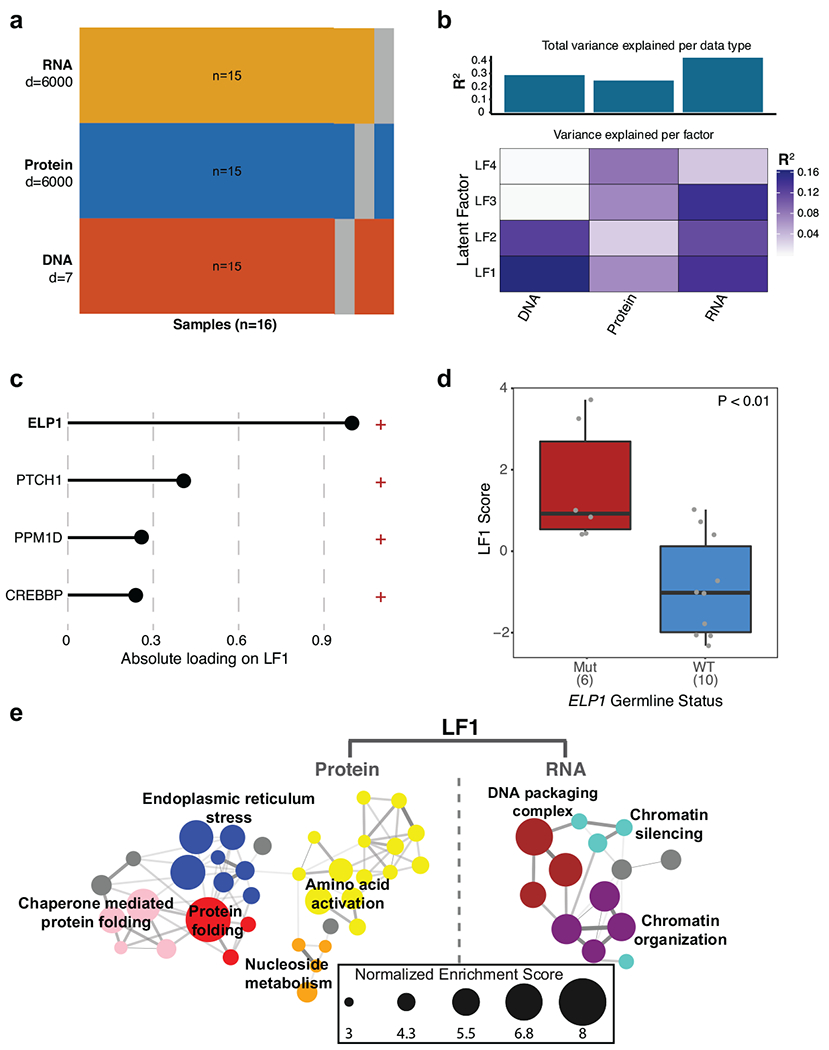
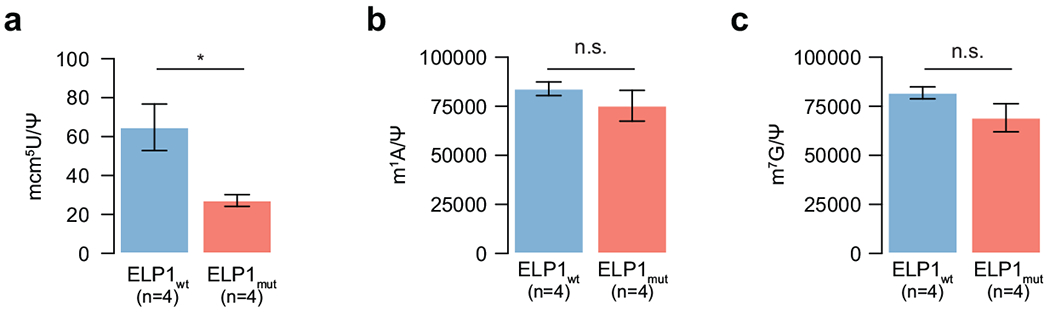
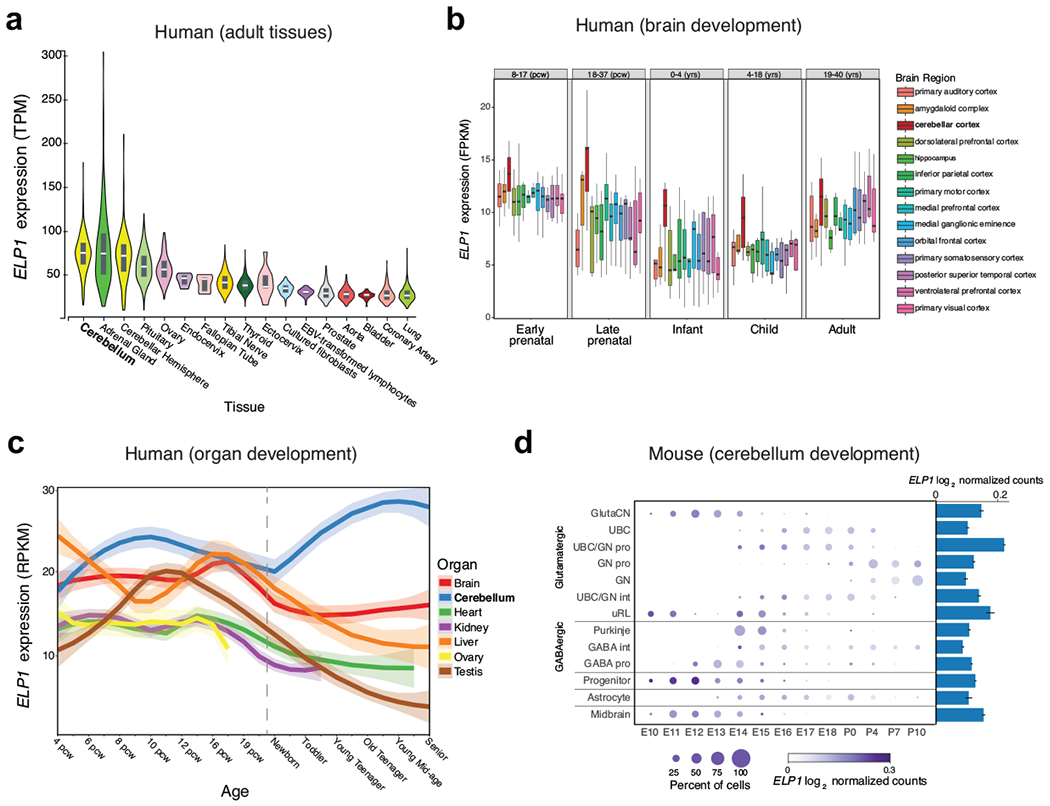
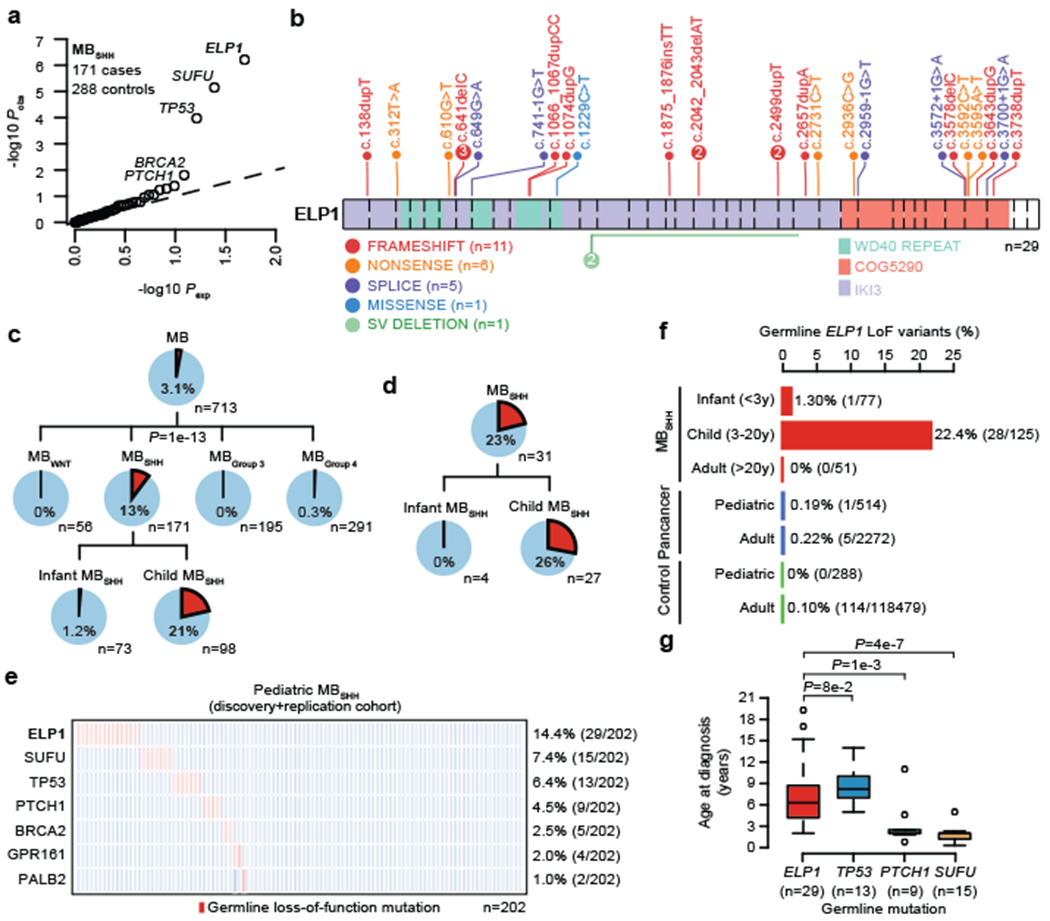
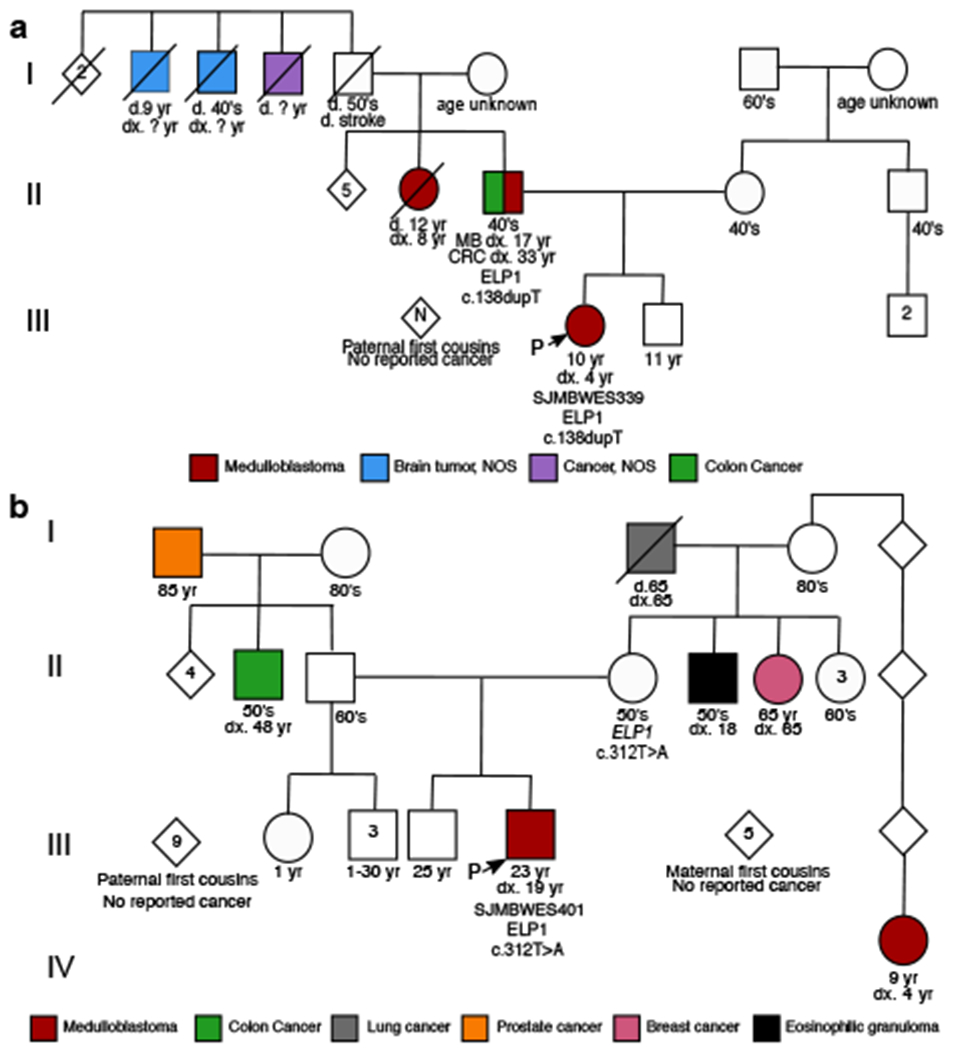
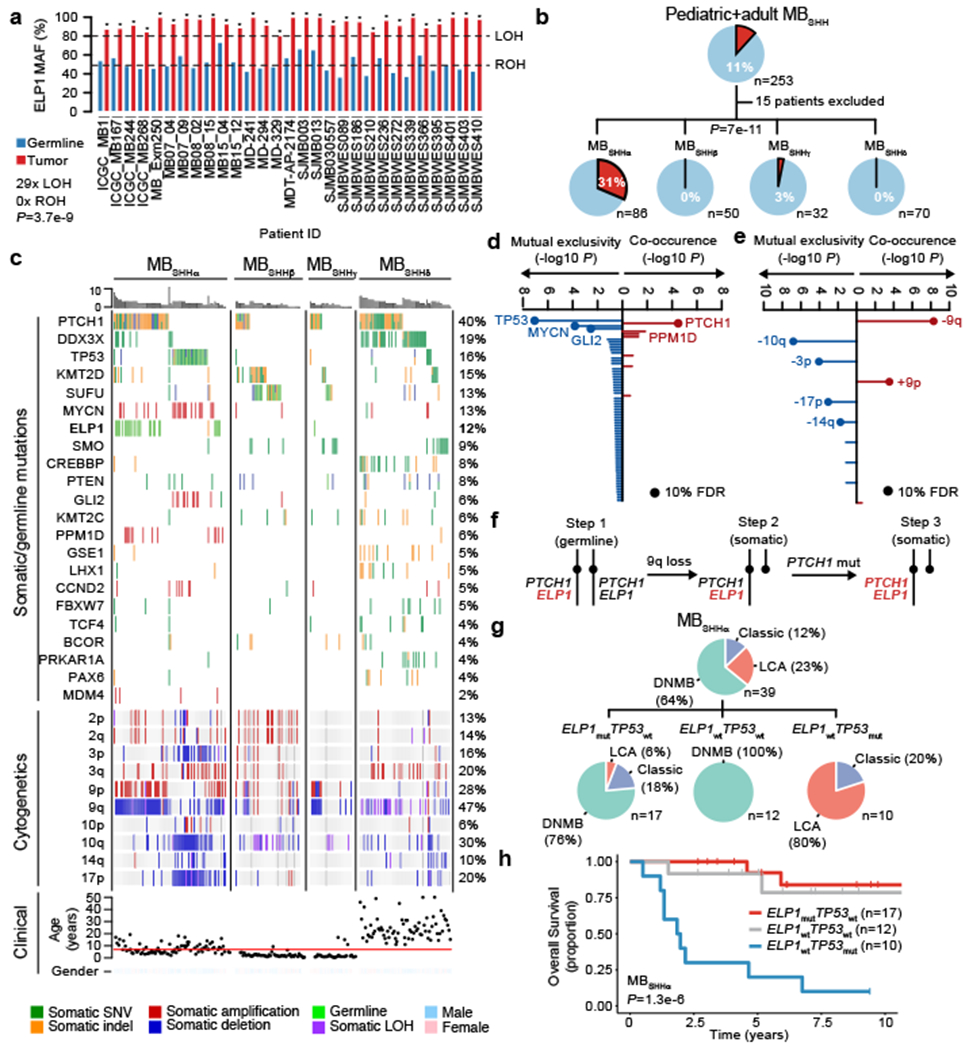
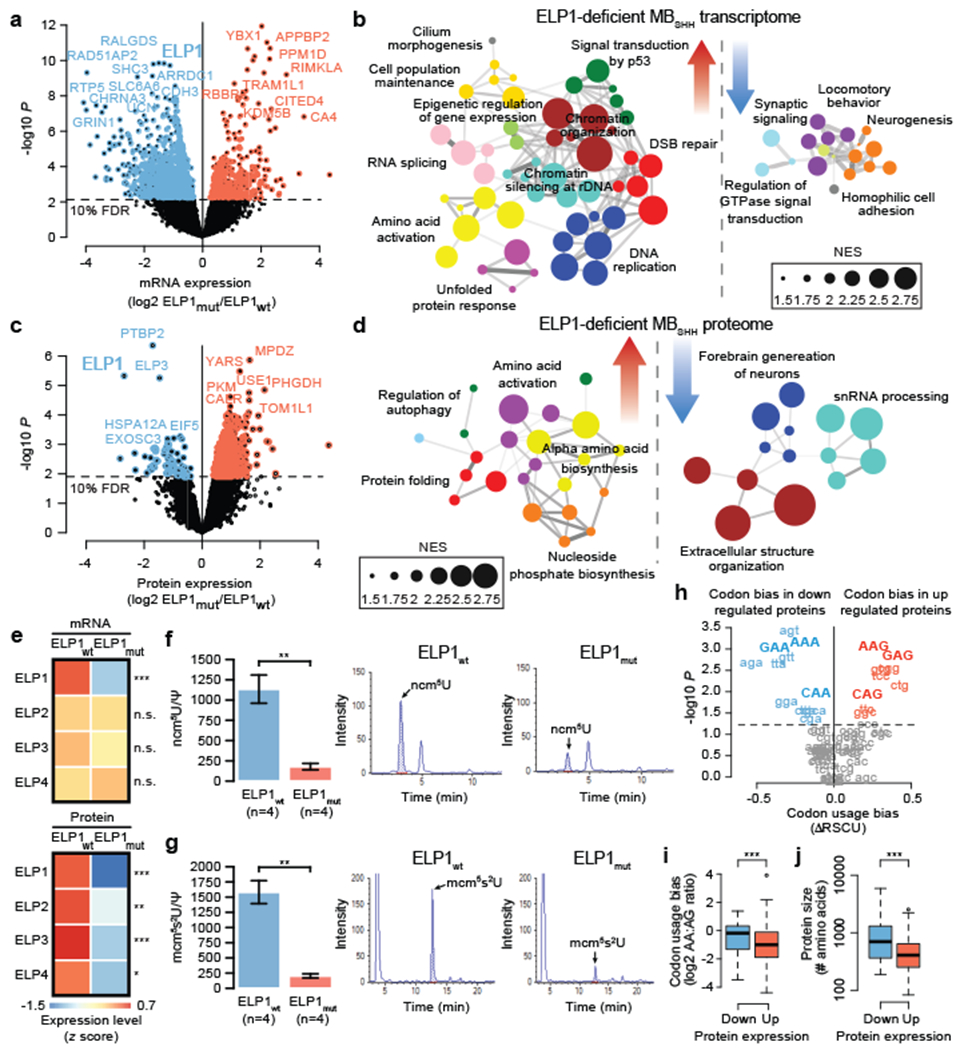
Cancer genomics has revealed many genes and core molecular processes that contribute to human malignancies, but the genetic and molecular bases of many rare cancers remains unclear. Genetic predisposition accounts for 5 to 10% of cancer diagnoses in children1,2, and genetic events that cooperate with known somatic driver events are poorly understood. Pathogenic germline variants in established cancer predisposition genes have been recently identified in 5% of patients with the malignant brain tumour medulloblastoma3. Here, by analysing all protein-coding genes, we identify and replicate rare germline loss-of-function variants across ELP1 in 14% of paediatric patients with the medulloblastoma subgroup Sonic Hedgehog (MBSHH). ELP1 was the most common medulloblastoma predisposition gene and increased the prevalence of genetic predisposition to 40% among paediatric patients with MBSHH. Parent-offspring and pedigree analyses identified two families with a history of paediatric medulloblastoma. ELP1-associated medulloblastomas were restricted to the molecular SHHα subtype4 and characterized by universal biallelic inactivation of ELP1 owing to somatic loss of chromosome arm 9q. Most ELP1-associated medulloblastomas also exhibited somatic alterations in PTCH1, which suggests that germline ELP1 loss-of-function variants predispose individuals to tumour development in combination with constitutive activation of SHH signalling. ELP1 is the largest subunit of the evolutionarily conserved Elongator complex, which catalyses translational elongation through tRNA modifications at the wobble (U34) position5,6. Tumours from patients with ELP1-associated MBSHH were characterized by a destabilized Elongator complex, loss of Elongator-dependent tRNA modifications, codon-dependent translational reprogramming, and induction of the unfolded protein response, consistent with loss of protein homeostasis due to Elongator deficiency in model systems7-9. Thus, genetic predisposition to proteome instability may be a determinant in the pathogenesis of paediatric brain cancers. These results support investigation of the role of protein homeostasis in other cancer types and potential for therapeutic interference.
Conflict of interest statement
Competing Interests
The authors declare no competing interests.
Figures
References
-
- Grobner SN, Worst BC, Weischenfeldt J, Buchhalter I, Kleinheinz K, Rudneva VA, Johann PD , Balasubramanian GP, Segura-Wang M, Brabetz S, Bender S, Hutter B, Sturm D, Pfaff E, Hubschmann D, Zipprich G, Heinold M, Eils J, Lawerenz C, Erkek S, Lambo S, Waszak S, Blattmann C, Borkhardt A, Kuhlen M, Eggert A, Fulda S, Gessler M, Wegert J, Kappler R, Baumhoer D, Burdach S, Kirschner-Schwabe R, Kontny U, Kulozik AE, Lohmann D, Hettmer S, Eckert C, Bielack S, Nathrath M, Niemeyer C, Richter GH, Schulte J, Siebert R, Westermann F, Molenaar JJ, Vassal G, Witt H, Project IP-S, Project IM-S, Burkhardt B, Kratz CP, Witt O, van Tilburg CM, Kramm CM, Fleischhack G, Dirksen U, Rutkowski S, Fruhwald M, von Hoff K, Wolf S, Klingebiel T, Koscielniak E, Landgraf P, Koster J, Resnick AC, Zhang J, Liu Y, Zhou X, Waanders AJ, Zwijnenburg DA, Raman P, Brors B, Weber UD, Northcott PA, Pajtler KW, Kool M, Piro RM, Korbel JO, Schlesner M, Eils R, Jones DTW, Lichter P, Chavez L, Zapatka M & Pfister SM The landscape of genomic alterations across childhood cancers. Nature 555, 321–327, doi:10.1038/nature25480 (2018). - DOI - PubMed
-
- Zhang J, Walsh MF, Wu G, Edmonson MN, Gruber TA, Easton J, Hedges D, Ma X, Zhou X, Yergeau DA, Wilkinson MR, Vadodaria B, Chen X, McGee RB, Hines-Dowell S, Nuccio R, Quinn E, Shurtleff SA, Rusch M, Patel A, Becksfort JB, Wang S, Weaver MS, Ding L, Mardis ER, Wilson RK, Gajjar A, Ellison DW, Pappo AS, Pui CH, Nichols KE & Downing JR Germline Mutations in Predisposition Genes in Pediatric Cancer. N Engl J Med 373, 2336–2346, doi:10.1056/NEJMoa1508054 (2015). - DOI - PMC - PubMed
-
- Waszak SM, Northcott PA, Buchhalter I, Robinson GW, Sutter C, Groebner S, Grand KB, Brugieres L, Jones DTW, Pajtler KW, Morrissy AS, Kool M, Sturm D, Chavez L, Ernst A, Brabetz S, Hain M, Zichner T, Segura-Wang M, Weischenfeldt J, Rausch T, Mardin BR, Zhou X, Baciu C, Lawerenz C, Chan JA, Varlet P, Guerrini-Rousseau L, Fults DW, Grajkowska W, Hauser P, Jabado N, Ra YS, Zitterbart K, Shringarpure SS, De La Vega FM, Bustamante CD, Ng HK, Perry A, MacDonald TJ, Hernaiz Driever P, Bendel AE, Bowers DC, McCowage G, Chintagumpala MM, Cohn R, Hassall T, Fleischhack G, Eggen T, Wesenberg F, Feychting M, Lannering B, Schuz J, Johansen C, Andersen TV, Roosli M, Kuehni CE, Grotzer M, Kjaerheim K , Monoranu CM, Archer TC, Duke E, Pomeroy SL, Shelagh R, Frank S, Sumerauer D, Scheurlen W, Ryzhova MV, Milde T, Kratz CP, Samuel D, Zhang J, Solomon DA, Marra M, Eils R, Bartram CR, von Hoff K, Rutkowski S, Ramaswamy V, Gilbertson RJ, Korshunov A, Taylor MD, Lichter P, Malkin D, Gajjar A, Korbel JO & Pfister SM Spectrum and prevalence of genetic predisposition in medulloblastoma: a retrospective genetic study and prospective validation in a clinical trial cohort. Lancet Oncol 19, 785–798, doi:10.1016/S1470-2045(18)30242-0 (2018). - DOI - PMC - PubMed
-
- Cavalli FMG, Remke M, Rampasek L, Peacock J, Shih DJH, Luu B, Garzia L, Torchia J, Nor C, Morrissy AS, Agnihotri S, Thompson YY, Kuzan-Fischer CM, Farooq H, Isaev K, Daniels C , Cho BK, Kim SK, Wang KC, Lee JY, Grajkowska WA, Perek-Polnik M, Vasiljevic A, Faure-Conter C, Jouvet A, Giannini C, Nageswara Rao AA, Li KKW, Ng HK, Eberhart CG, Pollack IF, Hamilton RL, Gillespie GY, Olson JM, Leary S, Weiss WA, Lach B, Chambless LB, Thompson RC, Cooper MK, Vibhakar R, Hauser P, van Veelen MC, Kros JM, French PJ , Ra YS, Kumabe T, Lopez-Aguilar E, Zitterbart K, Sterba J, Finocchiaro G, Massimino M, Van Meir EG, Osuka S, Shofuda T, Klekner A, Zollo M, Leonard JR, Rubin JB, Jabado N, Albrecht S, Mora J, Van Meter TE, Jung S, Moore AS, Hallahan AR, Chan JA, Tirapelli DPC, Carlotti CG, Fouladi M, Pimentel J, Faria CC, Saad AG, Massimi L, Liau LM, Wheeler H, Nakamura H, Elbabaa SK, Perezpena-Diazconti M, Chico Ponce de Leon F, Robinson S, Zapotocky M , Lassaletta A, Huang A, Hawkins CE, Tabori U, Bouffet E, Bartels U, Dirks PB., Rutka JT., Bader GD., Reimand J, Goldenberg A, Ramaswamy V & Taylor MD. Intertumoral Heterogeneity within Medulloblastoma Subgroups. Cancer Cell 31, 737–754 e736, doi:10.1016/j.ccell.2017.05.005 (2017). - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
