

Transcriptomic Analysis of Monocyte-Derived Non-Phagocytic Macrophages Favors a Role in Limiting Tissue Repair and Fibrosis
- PMID: 32296415
- PMCID: PMC7136412
- DOI: 10.3389/fimmu.2020.00405
Transcriptomic Analysis of Monocyte-Derived Non-Phagocytic Macrophages Favors a Role in Limiting Tissue Repair and Fibrosis
Erratum in
-
Corrigendum: Transcriptomic Analysis of Monocyte-Derived Non-Phagocytic Macrophages Favors a Role in Limiting Tissue Repair and Fibrosis.Front Immunol. 2020 May 20;11:1003. doi: 10.3389/fimmu.2020.01003. eCollection 2020. Front Immunol. 2020. PMID: 32508846 Free PMC article.
Abstract
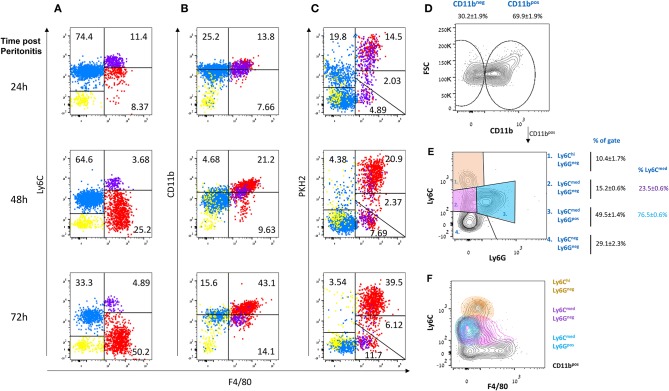
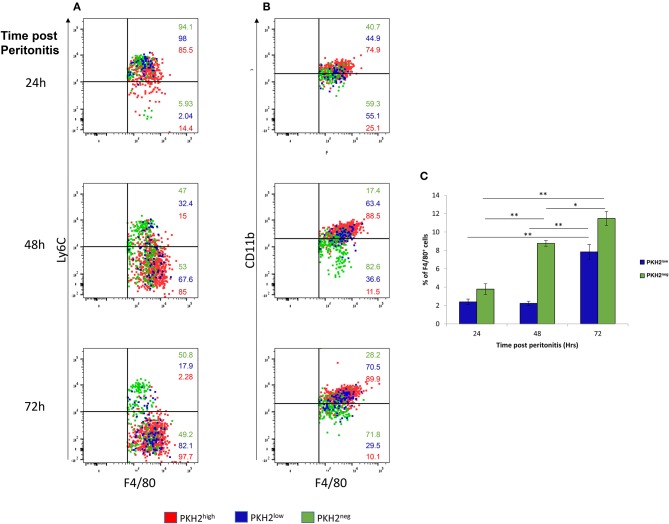
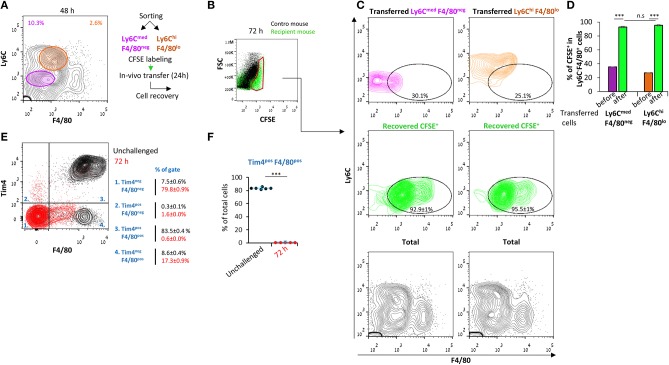
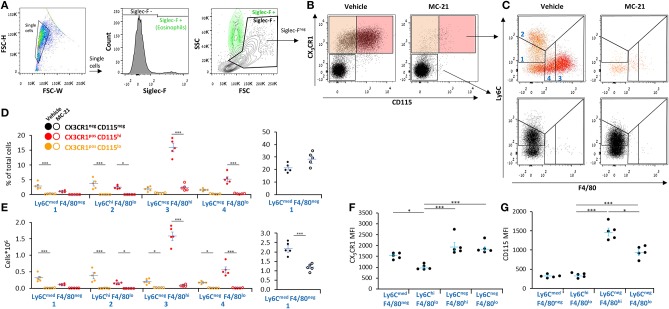
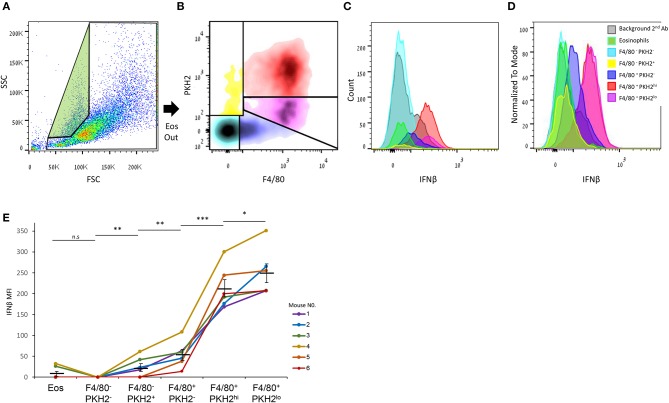
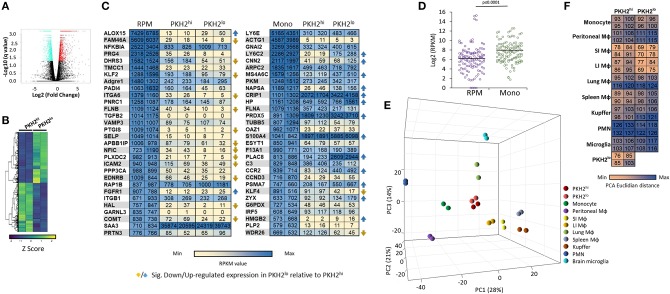
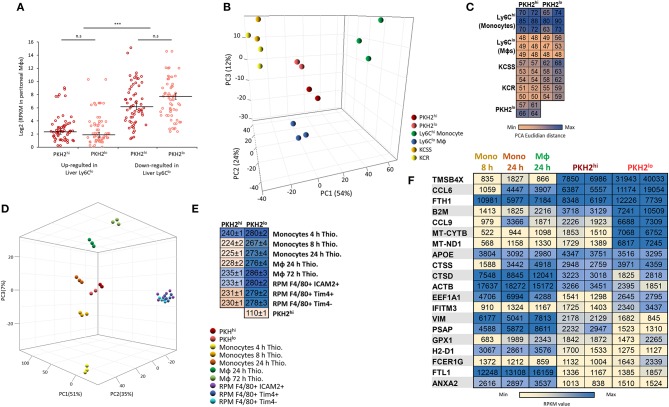
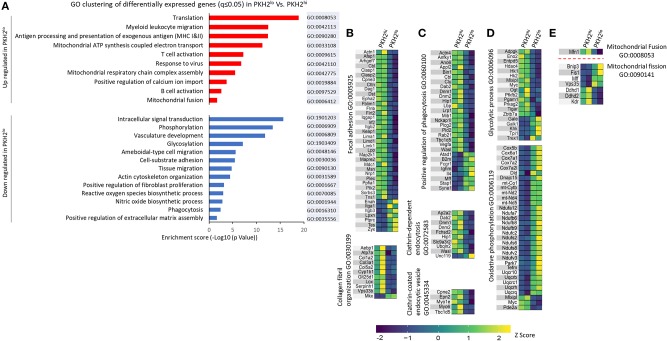
Monocyte-derived macrophages are readily differentiating cells that adapt their gene expression profile to environmental cues and functional needs. During the resolution of inflammation, monocytes initially differentiate to reparative phagocytic macrophages and later to pro-resolving non-phagocytic macrophages that produce high levels of IFNβ to boost resolutive events. Here, we performed in-depth analysis of phagocytic and non-phagocytic myeloid cells to reveal their distinct features. Unexpectedly, our analysis revealed that the non-phagocytic compartment of resolution phase myeloid cells is composed of Ly6CmedF4/80- and Ly6ChiF4/80lo monocytic cells in addition to the previously described Ly6C-F4/80+ satiated macrophages. In addition, we found that both Ly6C+ monocytic cells differentiate to Ly6C-F4/80+macrophages, and their migration to the peritoneum is CCR2 dependent. Notably, satiated macrophages expressed high levels of IFNβ, whereas non-phagocytic monocytes of either phenotype did not. A transcriptomic comparison of phagocytic and non-phagocytic resolution phase F4/80+ macrophages showed that both subtypes express similar gene signatures that make them distinct from other myeloid cells. Moreover, we confirmed that these macrophages express closer transcriptomes to monocytes than to resident peritoneal macrophages (RPM) and resemble resolutive Ly6Clo macrophages and monocyte-derived macrophages more than their precursors, inflammatory Ly6Chi monocytes, recovered following liver injury and healing, and thioglycolate-induced peritonitis, respectively. A direct comparison of these subsets indicated that the non-phagocytic transcriptome is dominated by satiated macrophages and downregulate gene clusters associated with excessive tissue repair and fibrosis, ROS and NO synthesis, glycolysis, and blood vessel morphogenesis. On the other hand, non-phagocytic macrophages enhance the expression of genes associated with migration, oxidative phosphorylation, and mitochondrial fission as well as anti-viral responses when compared to phagocytic macrophages. Notably, conversion from phagocytic to satiated macrophages is associated with a reduction in the expression of extracellular matrix constituents that were demonstrated to be associated with idiopathic pulmonary fibrosis (IPF). Thus, macrophage satiation during the resolution of inflammation seems to bring about a transcriptomic transition that resists tissue fibrosis and oxidative damage while promoting the restoration of tissue homeostasis to complete the resolution of inflammation.
Keywords: efferocytosis; fibrosis; inflammation; macrophages; transcriptional profiling.
Copyright © 2020 Butenko, Satyanarayanan, Assi, Schif-Zuck, Sher and Ariel.
Figures
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources