

Emerging Role of Compartmentalized G Protein-Coupled Receptor Signaling in the Cardiovascular Field
- PMID: 32296764
- PMCID: PMC7155194
- DOI: 10.1021/acsptsci.0c00006
Emerging Role of Compartmentalized G Protein-Coupled Receptor Signaling in the Cardiovascular Field
Abstract
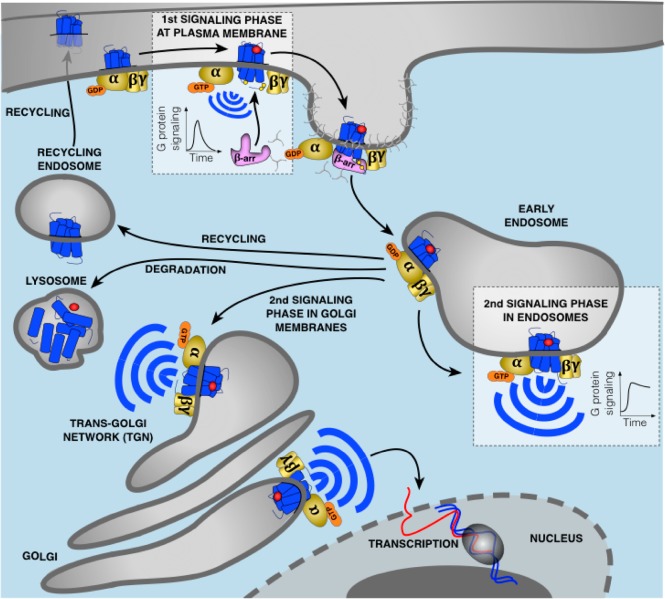
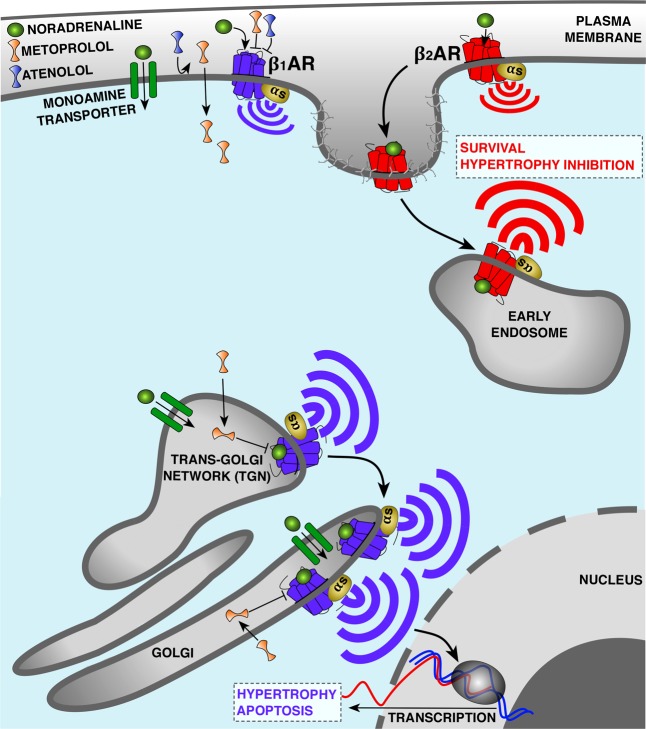
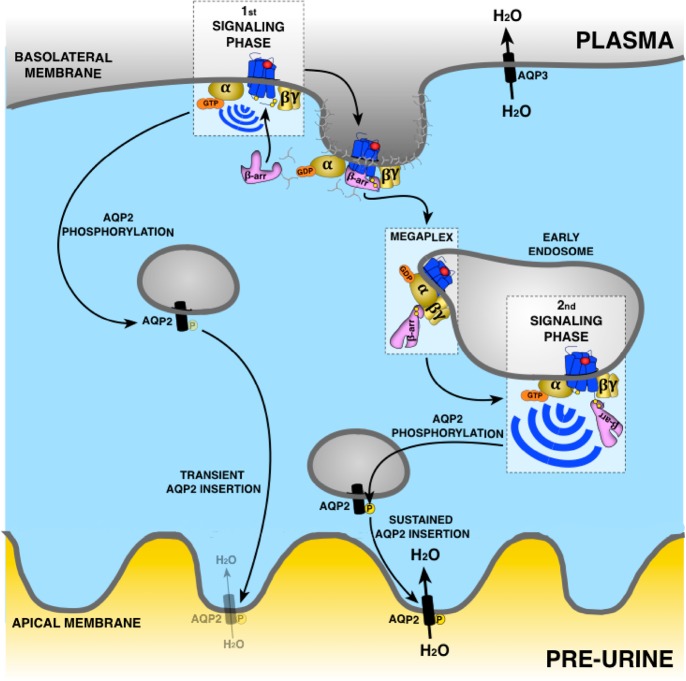
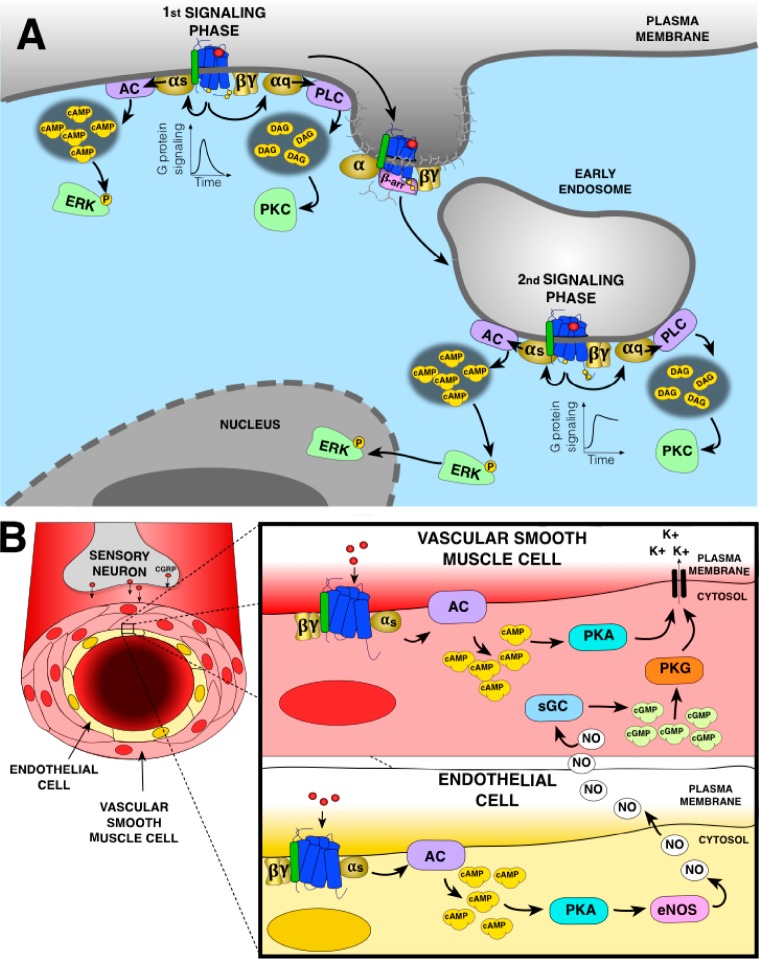
G protein-coupled receptors (GPCRs) are cell surface receptors that for many years have been considered to function exclusively at the plasma membrane, where they bind to extracellular ligands and activate G protein signaling cascades. According to the conventional model, these signaling events are rapidly terminated by β-arrestin (β-arr) recruitment to the activated GPCR resulting in signal desensitization and receptor internalization. However, during the past decade, emerging evidence suggest that many GPCRs can continue to activate G proteins from intracellular compartments after they have been internalized. G protein signaling from intracellular compartments is in general more sustained compared to G protein signaling at the plasma membrane. Notably, the particular location closer to the nucleus is beneficial for selective cellular functions such as regulation of gene transcription. Here, we review key GPCRs that undergo compartmentalized G protein signaling and discuss molecular considerations and requirements for this signaling to occur. Our main focus will be on receptors involved in the regulation of important physiological and pathological cardiovascular functions. We also discuss how sustained G protein activation from intracellular compartments may be involved in cellular functions that are distinct from functions regulated by plasma membrane G protein signaling, and the corresponding significance in cardiovascular physiology.
Copyright © 2020 American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous