

Mechanisms generating cancer genome complexity from a single cell division error
- PMID: 32299917
- PMCID: PMC7347108
- DOI: 10.1126/science.aba0712
Mechanisms generating cancer genome complexity from a single cell division error
Abstract
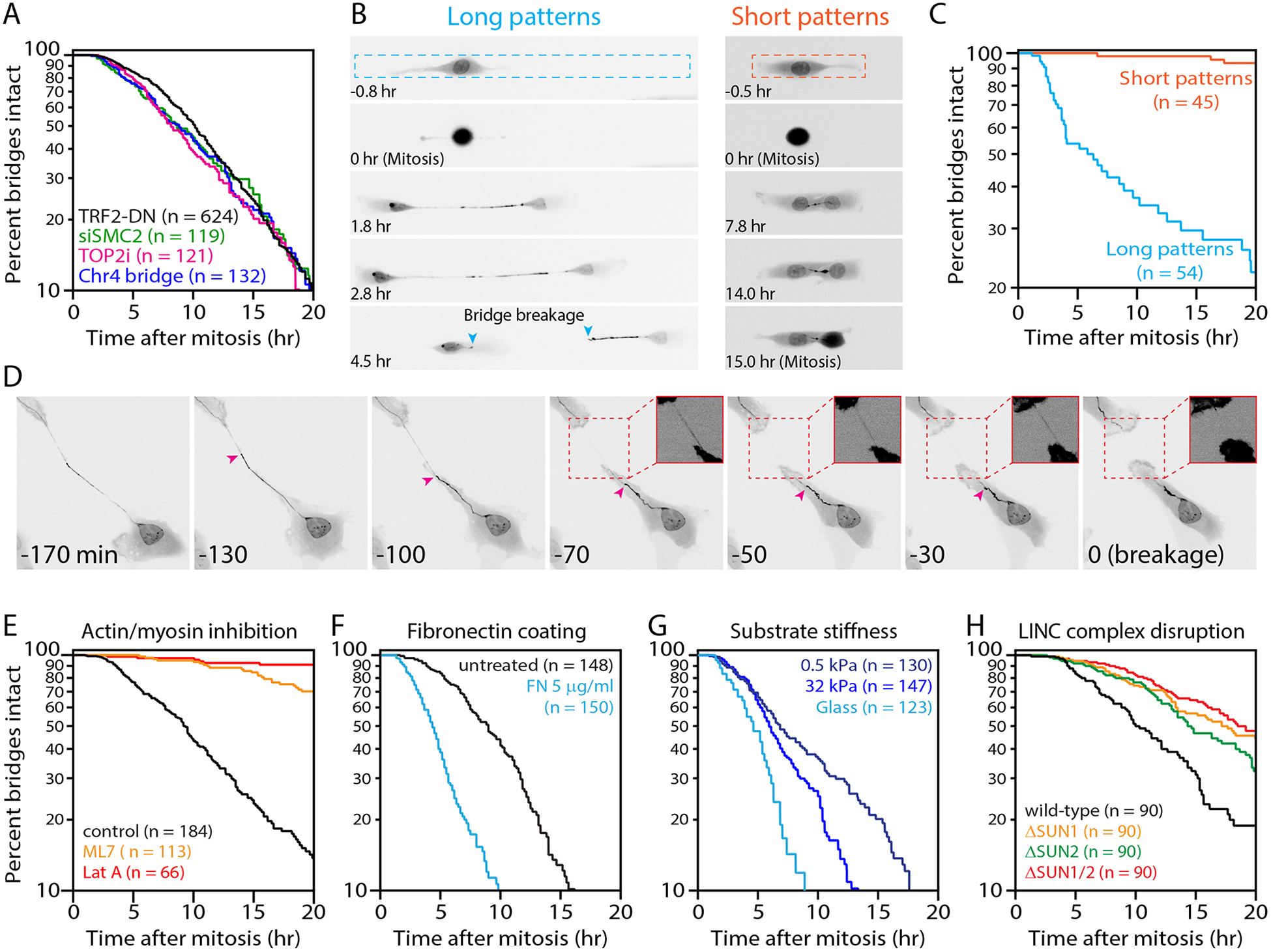
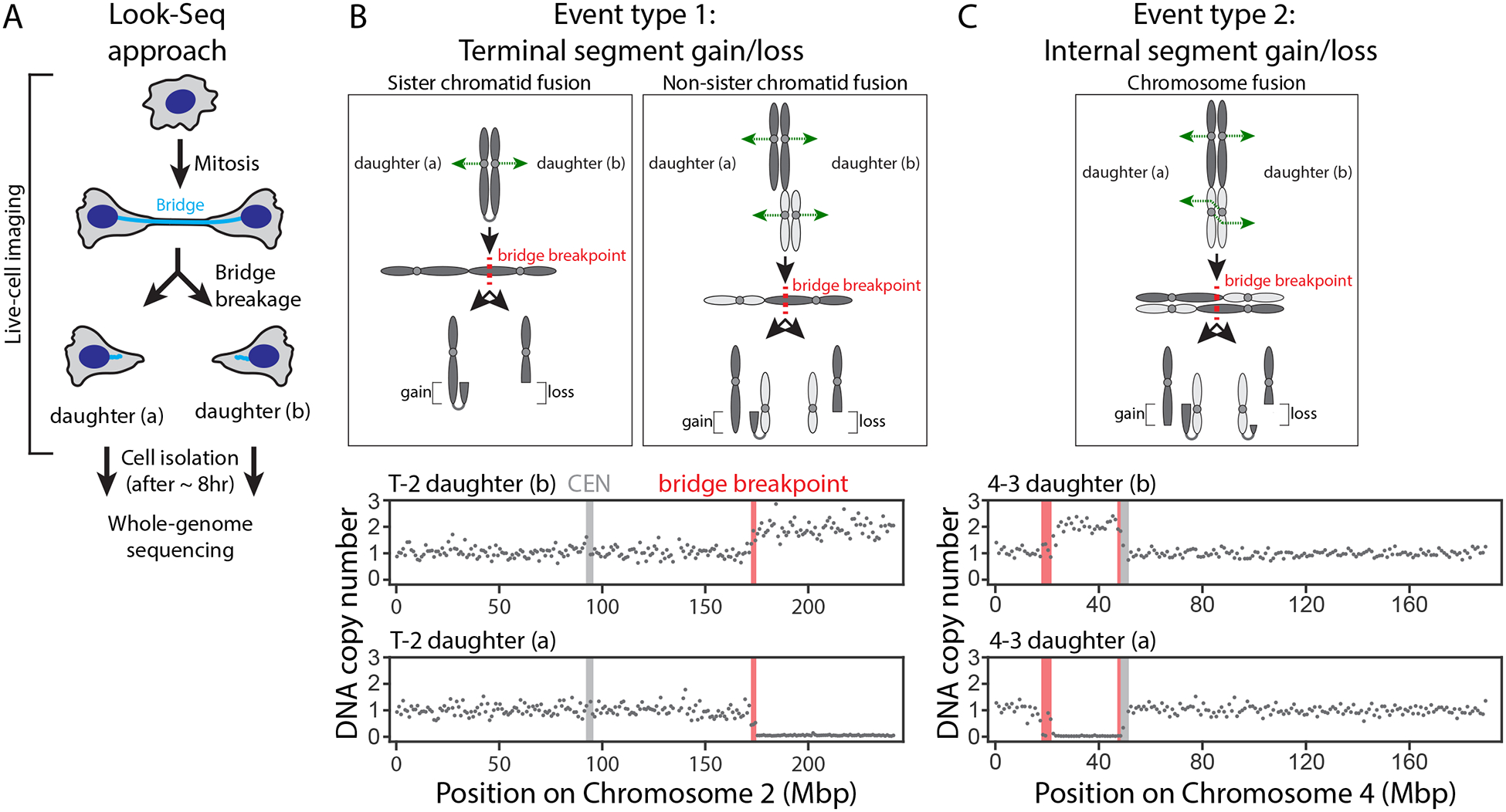
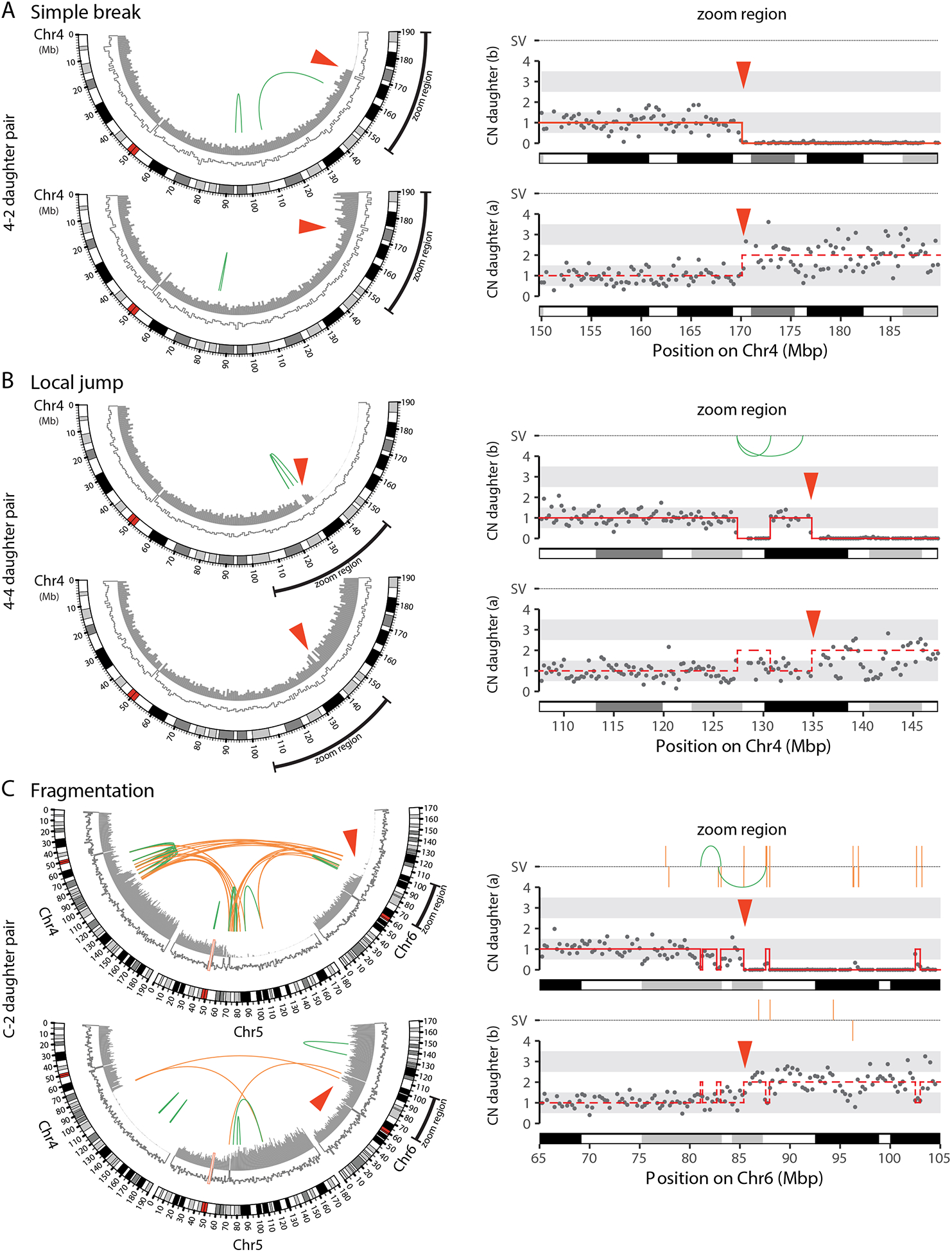
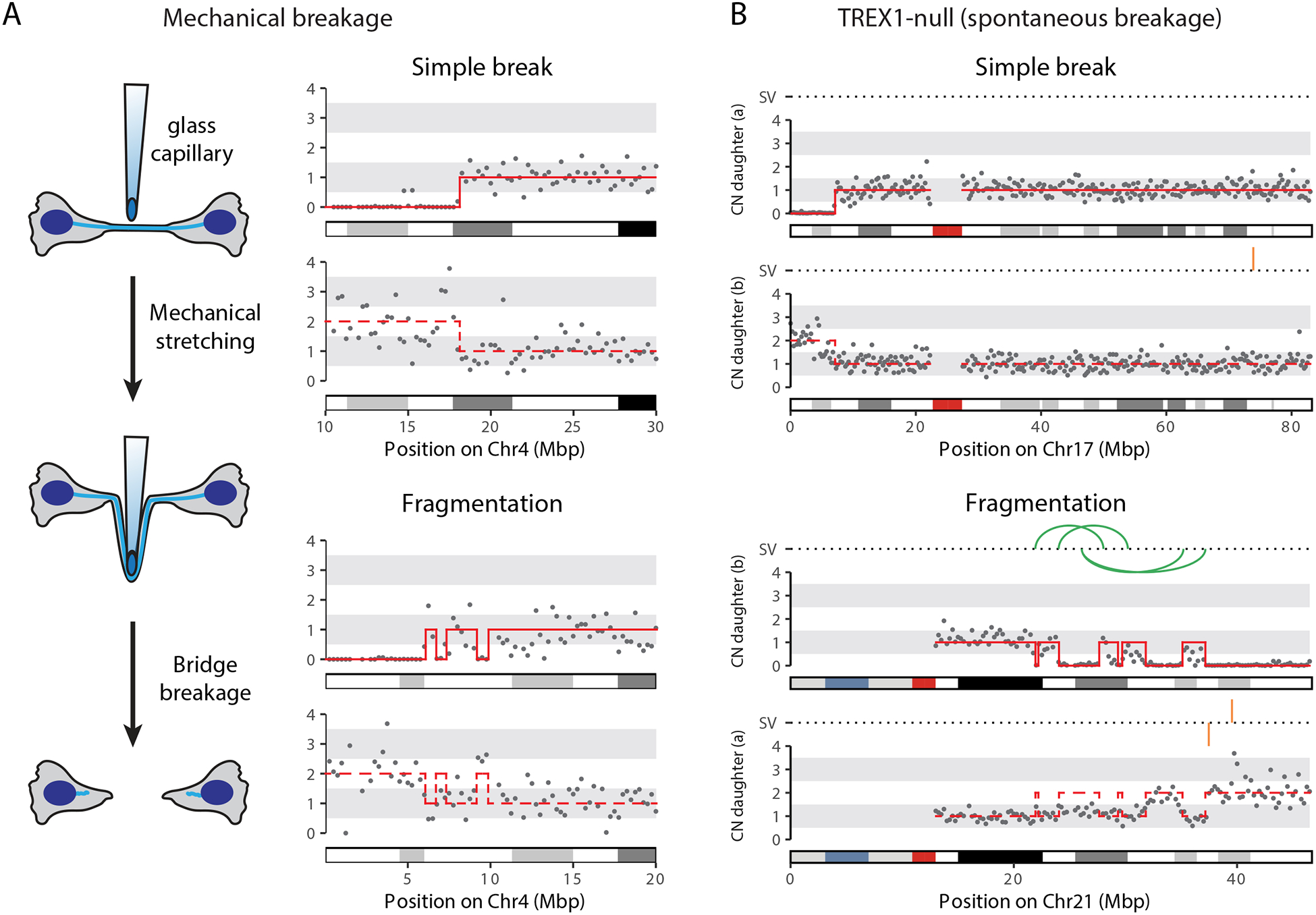
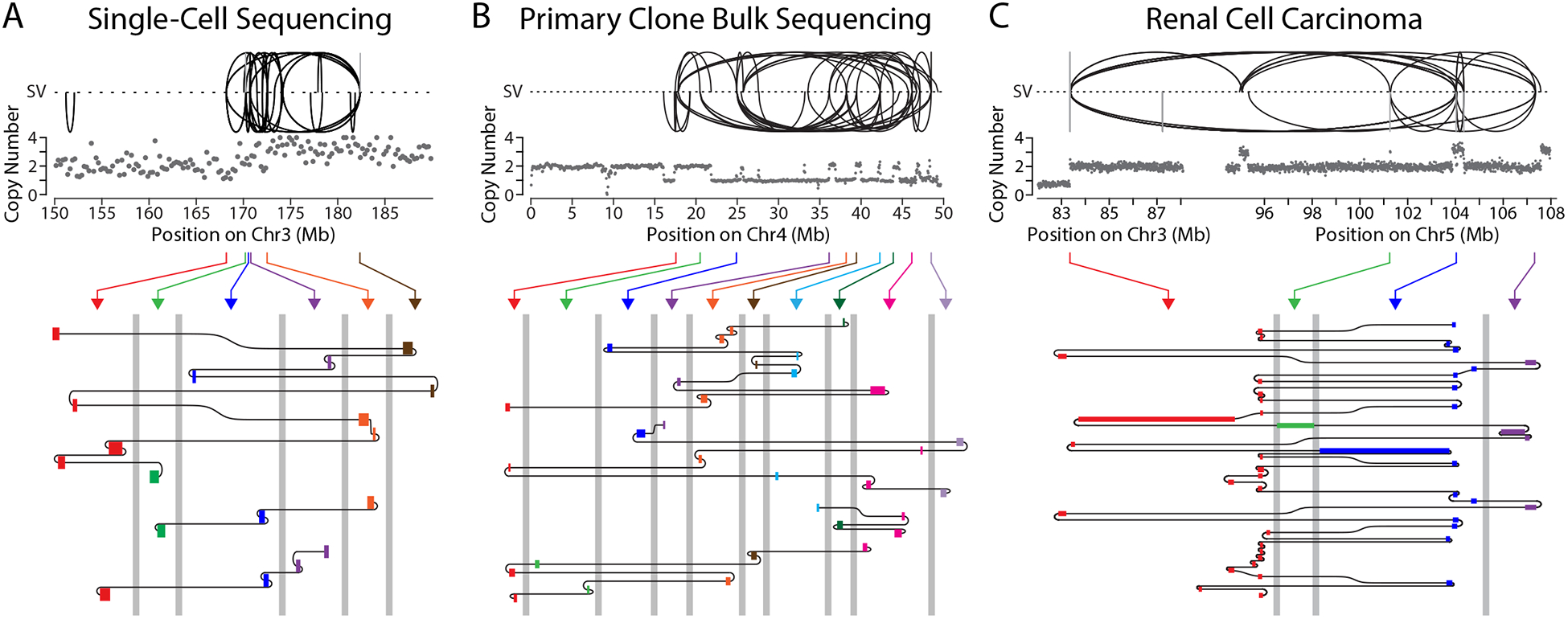
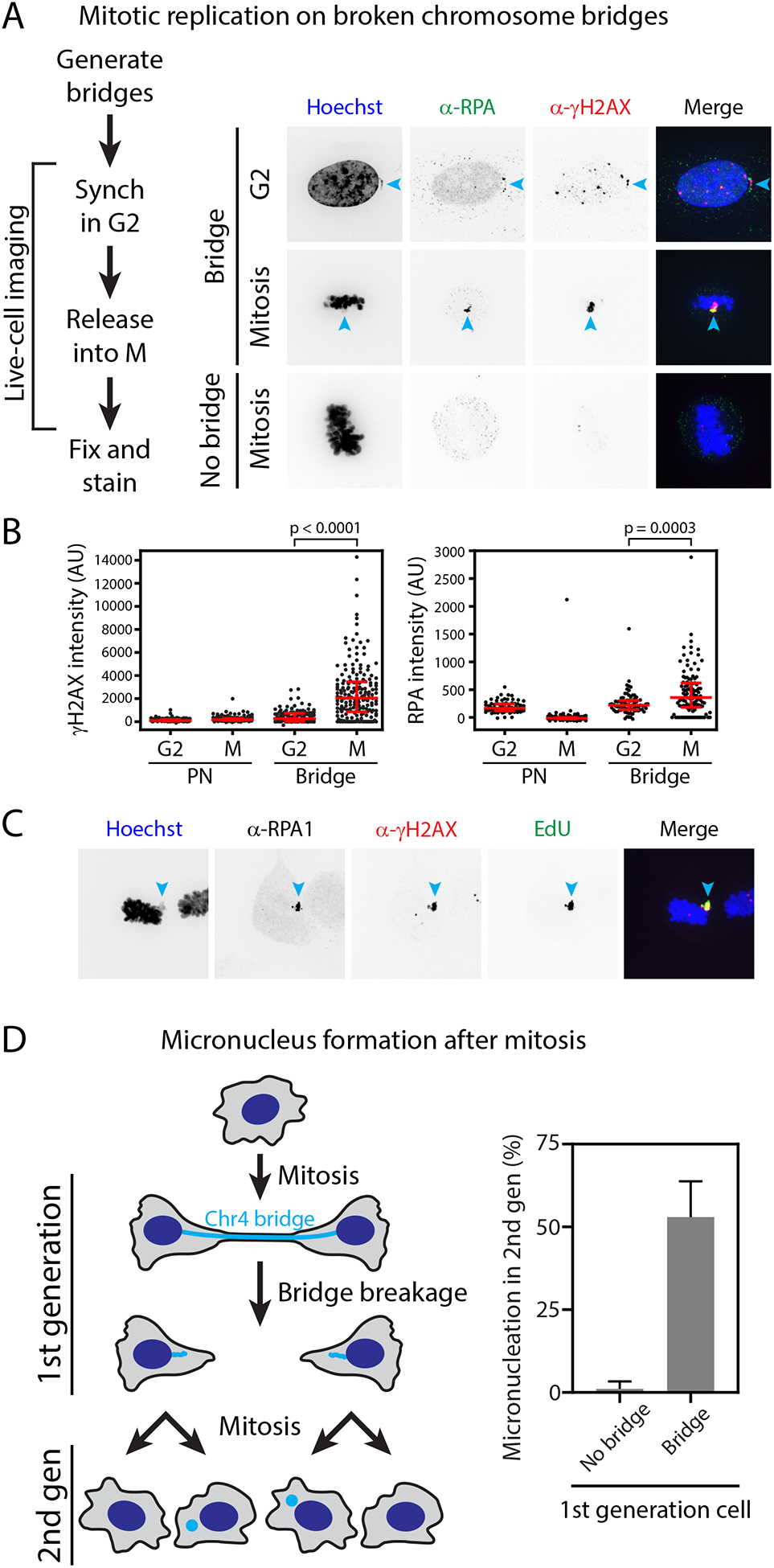
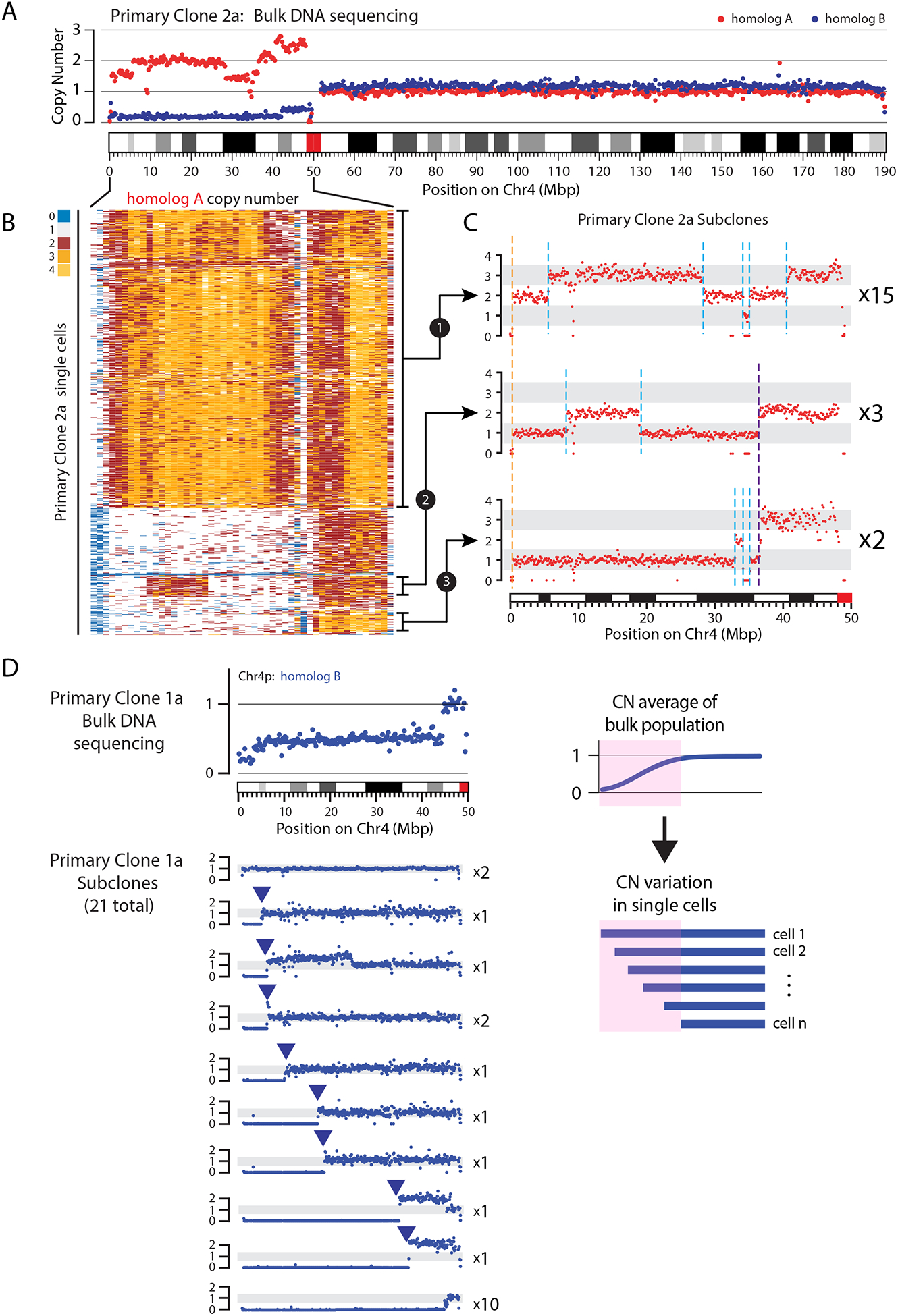
The chromosome breakage-fusion-bridge (BFB) cycle is a mutational process that produces gene amplification and genome instability. Signatures of BFB cycles can be observed in cancer genomes alongside chromothripsis, another catastrophic mutational phenomenon. We explain this association by elucidating a mutational cascade that is triggered by a single cell division error-chromosome bridge formation-that rapidly increases genomic complexity. We show that actomyosin forces are required for initial bridge breakage. Chromothripsis accumulates, beginning with aberrant interphase replication of bridge DNA. A subsequent burst of DNA replication in the next mitosis generates extensive DNA damage. During this second cell division, broken bridge chromosomes frequently missegregate and form micronuclei, promoting additional chromothripsis. We propose that iterations of this mutational cascade generate the continuing evolution and subclonal heterogeneity characteristic of many human cancers.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
COMPETING INTERESTS
The authors declare no competing interests.
Figures
Comment in
-
Burning bridges in cancer genomes.Science. 2020 Apr 17;368(6488):240-241. doi: 10.1126/science.abb4899. Science. 2020. PMID: 32299935 No abstract available.
References
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
