

Metabolomics and mass spectrometry imaging reveal channeled de novo purine synthesis in cells
- PMID: 32299949
- PMCID: PMC7494208
- DOI: 10.1126/science.aaz6465
Metabolomics and mass spectrometry imaging reveal channeled de novo purine synthesis in cells
Abstract
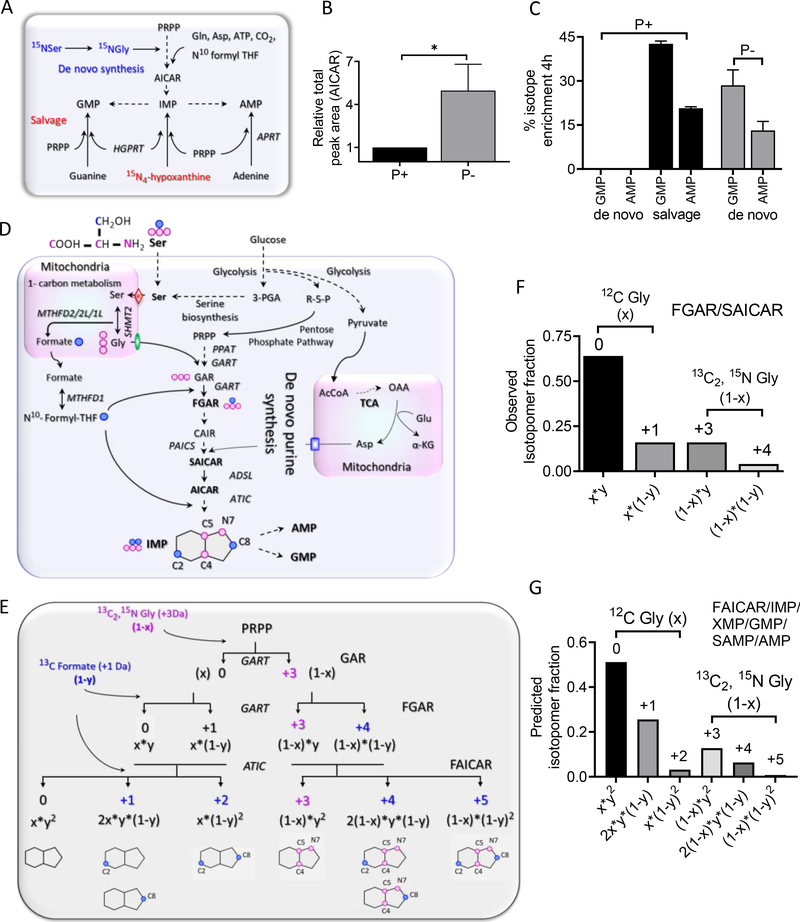
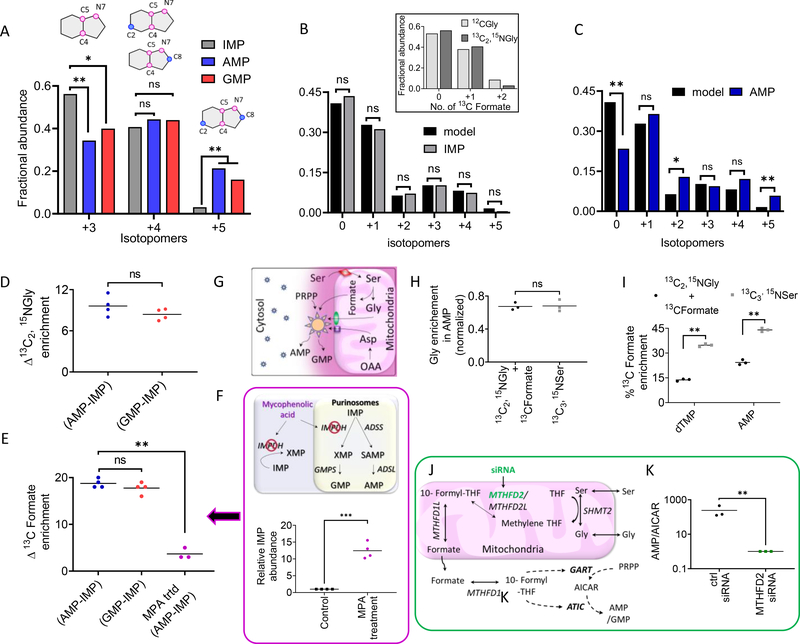
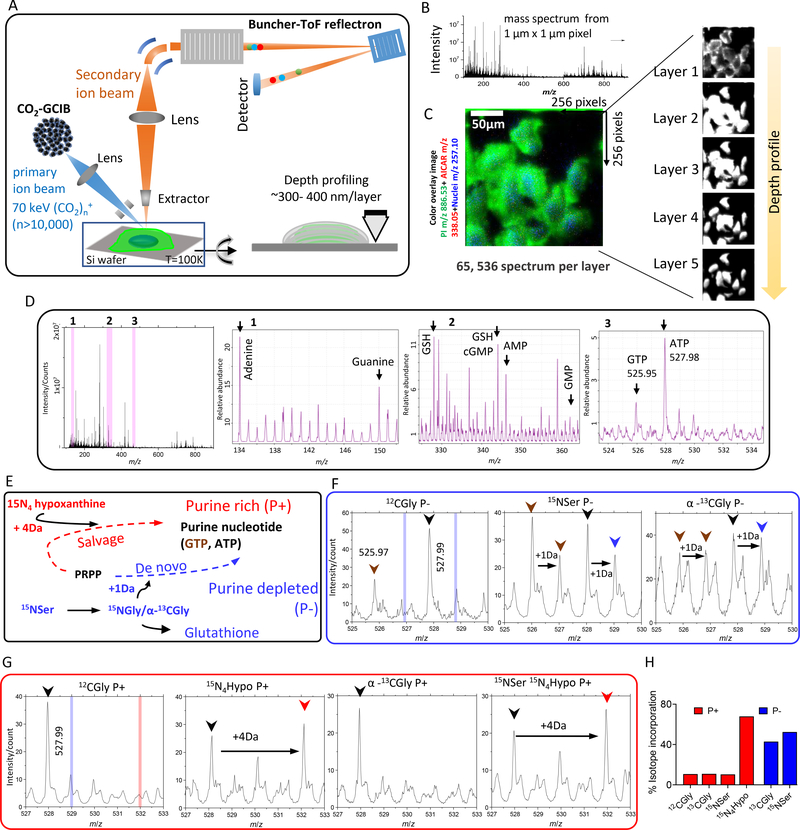
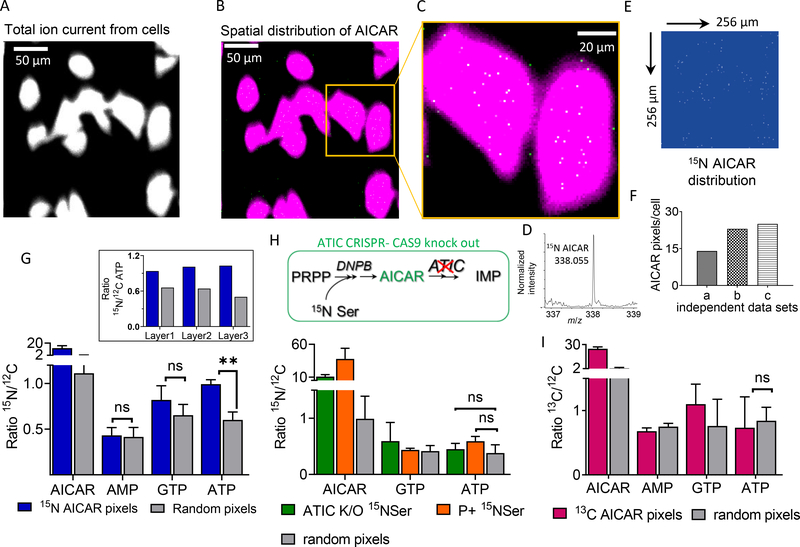
Metabolons, multiprotein complexes consisting of sequential enzymes of a metabolic pathway, are proposed to be biosynthetic "hotspots" within the cell. However, experimental demonstration of their presence and functions has remained challenging. We used metabolomics and in situ three-dimensional submicrometer chemical imaging of single cells by gas cluster ion beam secondary ion mass spectrometry (GCIB-SIMS) to directly visualize de novo purine biosynthesis by a multienzyme complex, the purinosome. We found that purinosomes comprise nine enzymes that act synergistically, channeling the pathway intermediates to synthesize purine nucleotides, increasing the pathway flux, and influencing the adenosine monophosphate/guanosine monophosphate ratio. Our work also highlights the application of high-resolution GCIB-SIMS for multiplexed biomolecular analysis at the level of single cells.
Copyright © 2020 The Authors, some rights reserved; exclusive licensee American Association for the Advancement of Science. No claim to original U.S. Government Works.
Conflict of interest statement
Figures
Comment in
-
Probing metabolism in time and space.Science. 2020 Apr 17;368(6488):241-242. doi: 10.1126/science.abb3094. Science. 2020. PMID: 32299936 No abstract available.
References
-
- Buchanan JM, Hartman SC, in Advances in Enzymology and Related Areas of Molecular Biology, Vol. 21, Nord FF, Ed. (Wiley, 1959), pp. 199–261.
-
- Hartman SC, Buchanan JM, Annu. Rev. Biochem. 28, 365–410 (1959). - PubMed
-
- Greenberg GR, Jaenicke L, in CIBA Foundation Symposium on the Chemistry and Biology Of Purines, Wolstenholme GEW, O’Connor CM, Eds. (Wiley, 1957), pp. 204–232.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
