

Recapitulation of the accessible interface of biopsy-derived canine intestinal organoids to study epithelial-luminal interactions
- PMID: 32302323
- PMCID: PMC7164685
- DOI: 10.1371/journal.pone.0231423
Recapitulation of the accessible interface of biopsy-derived canine intestinal organoids to study epithelial-luminal interactions
Abstract
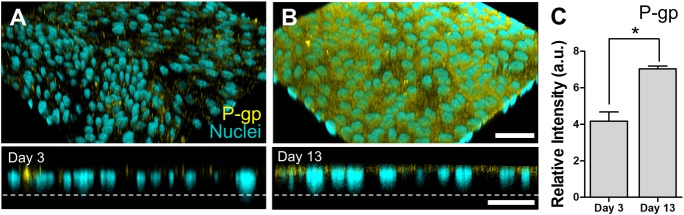
Recent advances in canine intestinal organoids have expanded the option for building a better in vitro model to investigate translational science of intestinal physiology and pathology between humans and animals. However, the three-dimensional geometry and the enclosed lumen of canine intestinal organoids considerably hinder the access to the apical side of epithelium for investigating the nutrient and drug absorption, host-microbiome crosstalk, and pharmaceutical toxicity testing. Thus, the creation of a polarized epithelial interface accessible from apical or basolateral side is critical. Here, we demonstrated the generation of an intestinal epithelial monolayer using canine biopsy-derived colonic organoids (colonoids). We optimized the culture condition to form an intact monolayer of the canine colonic epithelium on a nanoporous membrane insert using the canine colonoids over 14 days. Transmission and scanning electron microscopy revealed a physiological brush border interface covered by the microvilli with glycocalyx, as well as the presence of mucin granules, tight junctions, and desmosomes. The population of stem cells as well as differentiated lineage-dependent epithelial cells were verified by immunofluorescence staining and RNA in situ hybridization. The polarized expression of P-glycoprotein efflux pump was confirmed at the apical membrane. Also, the epithelial monolayer formed tight- and adherence-junctional barrier within 4 days, where the transepithelial electrical resistance and apparent permeability were inversely correlated. Hence, we verified the stable creation, maintenance, differentiation, and physiological function of a canine intestinal epithelial barrier, which can be useful for pharmaceutical and biomedical researches.
Conflict of interest statement
J.P.M., A.E.J., K.A., and H.J.K. are co-founders of 3D Health Solutions Inc. and hold an equity interest in the company. T.A. is employed by and receives a salary from 3D Health Solutions Inc. This does not alter our adherence to PLOS ONE policies on sharing data and materials.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
