

Role of MPK4 in pathogen-associated molecular pattern-triggered alternative splicing in Arabidopsis
- PMID: 32302366
- PMCID: PMC7164602
- DOI: 10.1371/journal.ppat.1008401
Role of MPK4 in pathogen-associated molecular pattern-triggered alternative splicing in Arabidopsis
Abstract
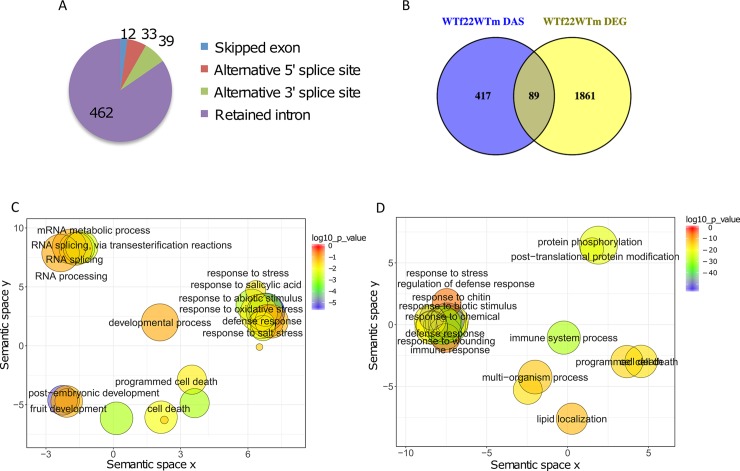
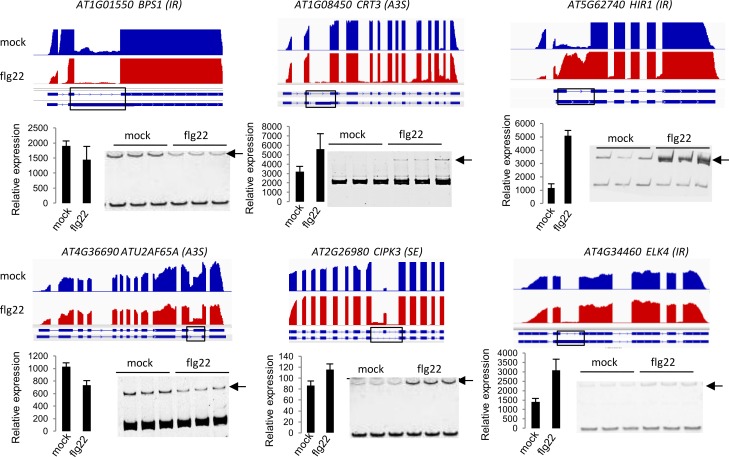
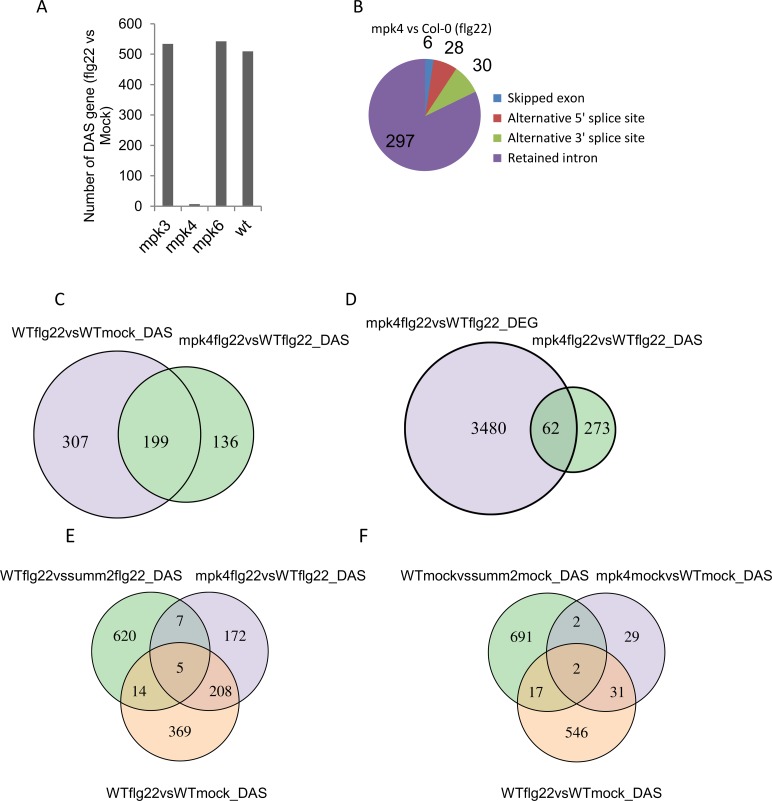
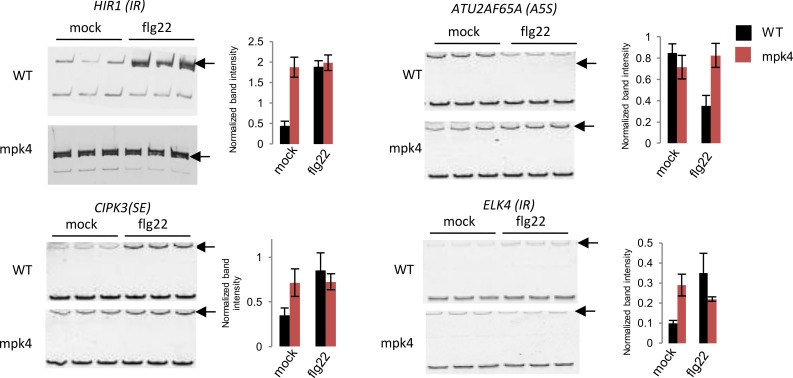
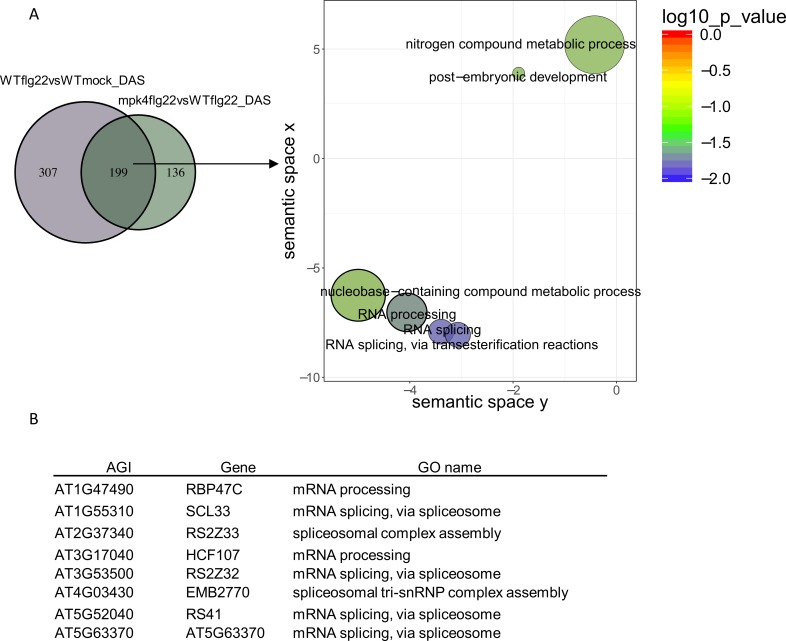
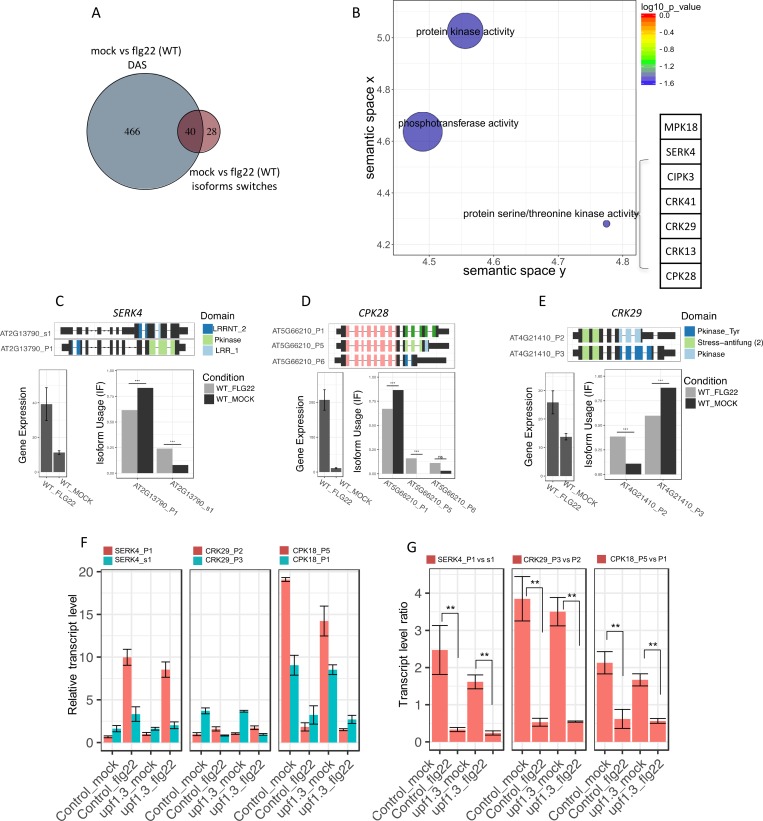
Alternative splicing (AS) of pre-mRNAs in plants is an important mechanism of gene regulation in environmental stress tolerance but plant signals involved are essentially unknown. Pathogen-associated molecular pattern (PAMP)-triggered immunity (PTI) is mediated by mitogen-activated protein kinases and the majority of PTI defense genes are regulated by MPK3, MPK4 and MPK6. These responses have been mainly analyzed at the transcriptional level, however many splicing factors are direct targets of MAPKs. Here, we studied alternative splicing induced by the PAMP flagellin in Arabidopsis. We identified 506 PAMP-induced differentially alternatively spliced (DAS) genes. Importantly, of the 506 PAMP-induced DAS genes, only 89 overlap with the set of 1950 PAMP-induced differentially expressed genes (DEG), indicating that transcriptome analysis does not identify most DAS events. Global DAS analysis of mpk3, mpk4, and mpk6 mutants in the absence of PAMP treatment showed no major splicing changes. However, in contrast to MPK3 and MPK6, MPK4 was found to be a key regulator of PAMP-induced DAS events as the AS of a number of splicing factors and immunity-related protein kinases is affected, such as the calcium-dependent protein kinase CPK28, the cysteine-rich receptor like kinases CRK13 and CRK29 or the FLS2 co-receptor SERK4/BKK1. Although MPK4 is guarded by SUMM2 and consequently, the mpk4 dwarf and DEG phenotypes are suppressed in mpk4 summ2 mutants, MPK4-dependent DAS is not suppressed by SUMM2, supporting the notion that PAMP-triggered MPK4 activation mediates regulation of alternative splicing.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
