

Competing Protein-RNA Interaction Networks Control Multiphase Intracellular Organization
- PMID: 32302570
- PMCID: PMC7816278
- DOI: 10.1016/j.cell.2020.03.050
Competing Protein-RNA Interaction Networks Control Multiphase Intracellular Organization
Abstract
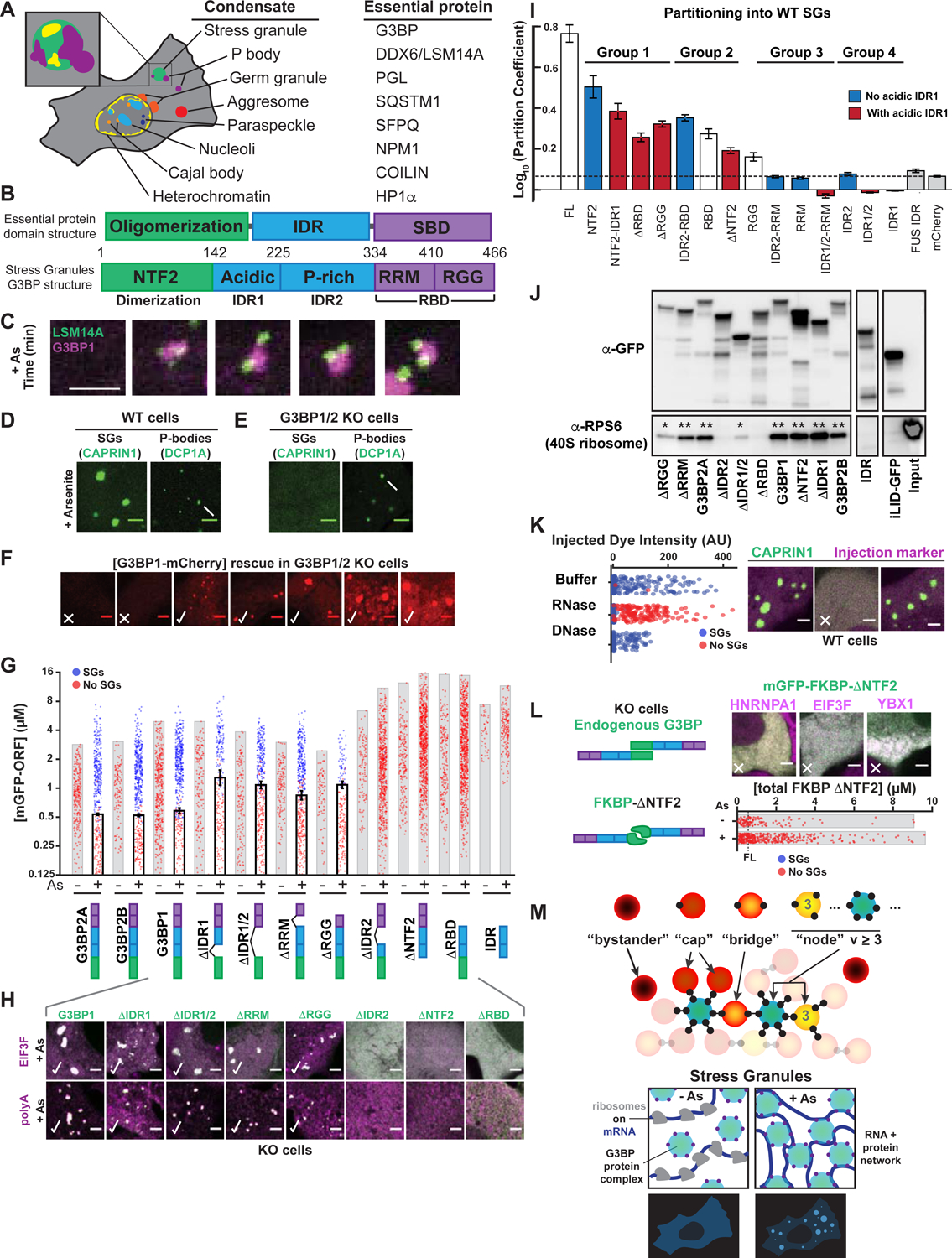
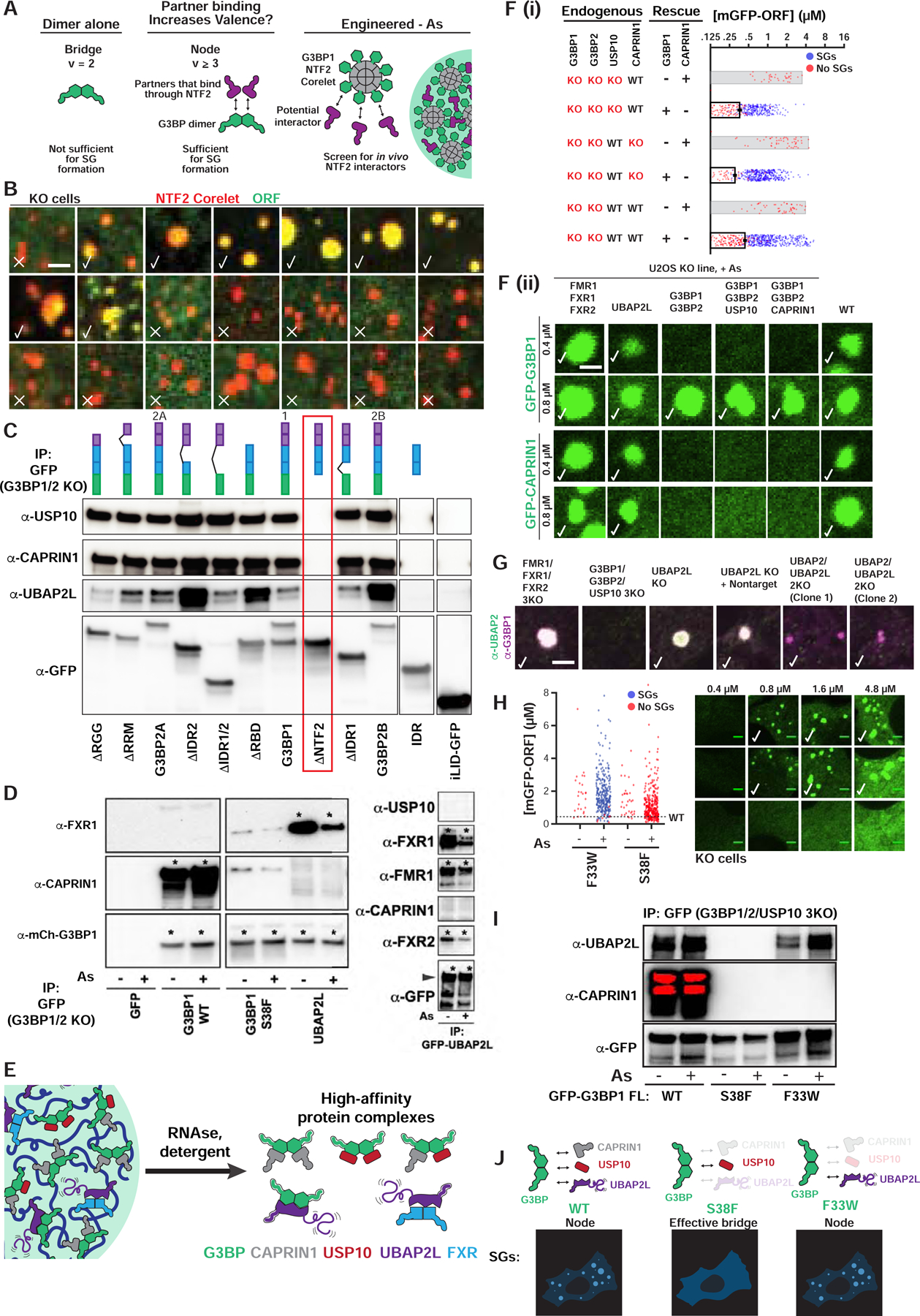
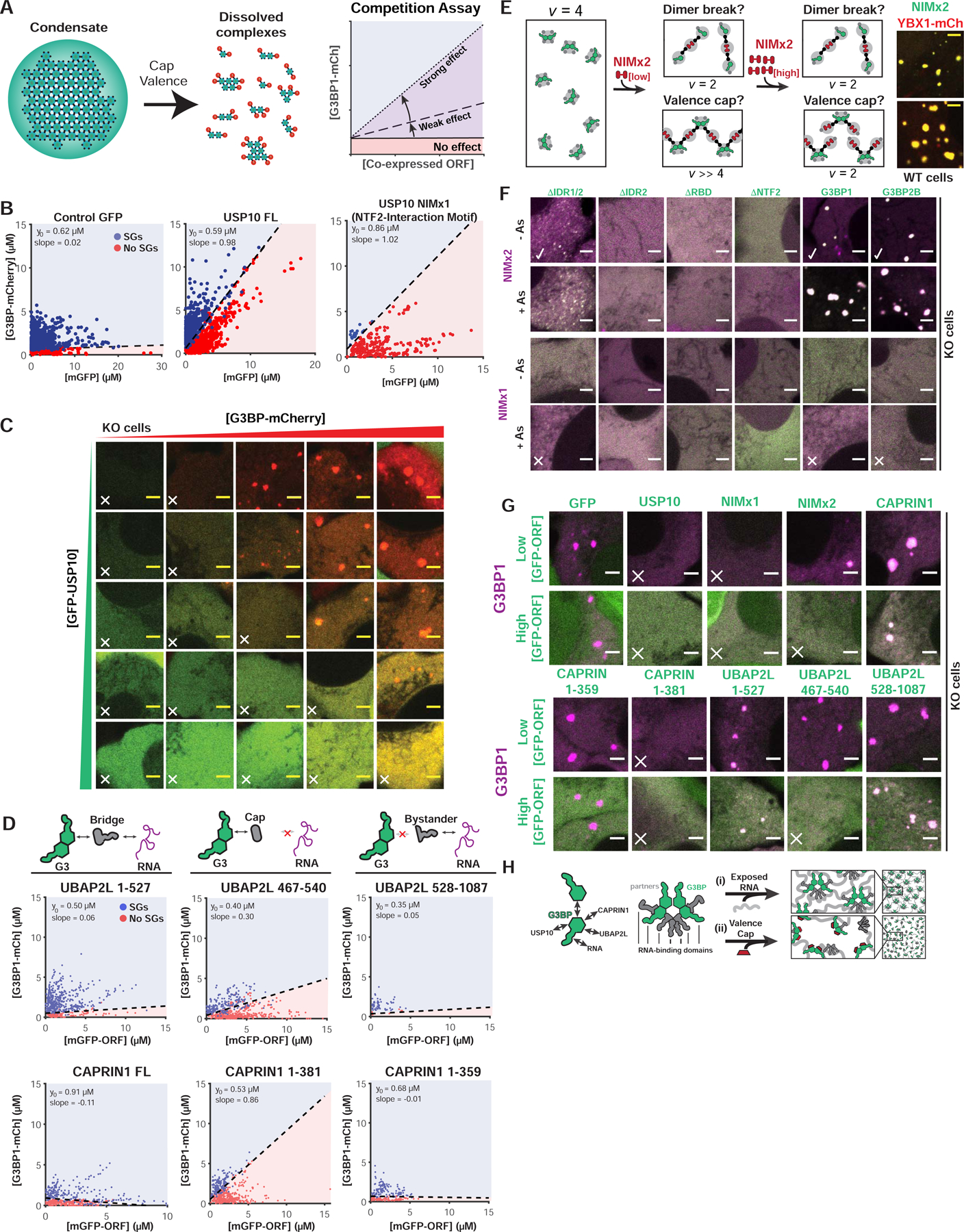
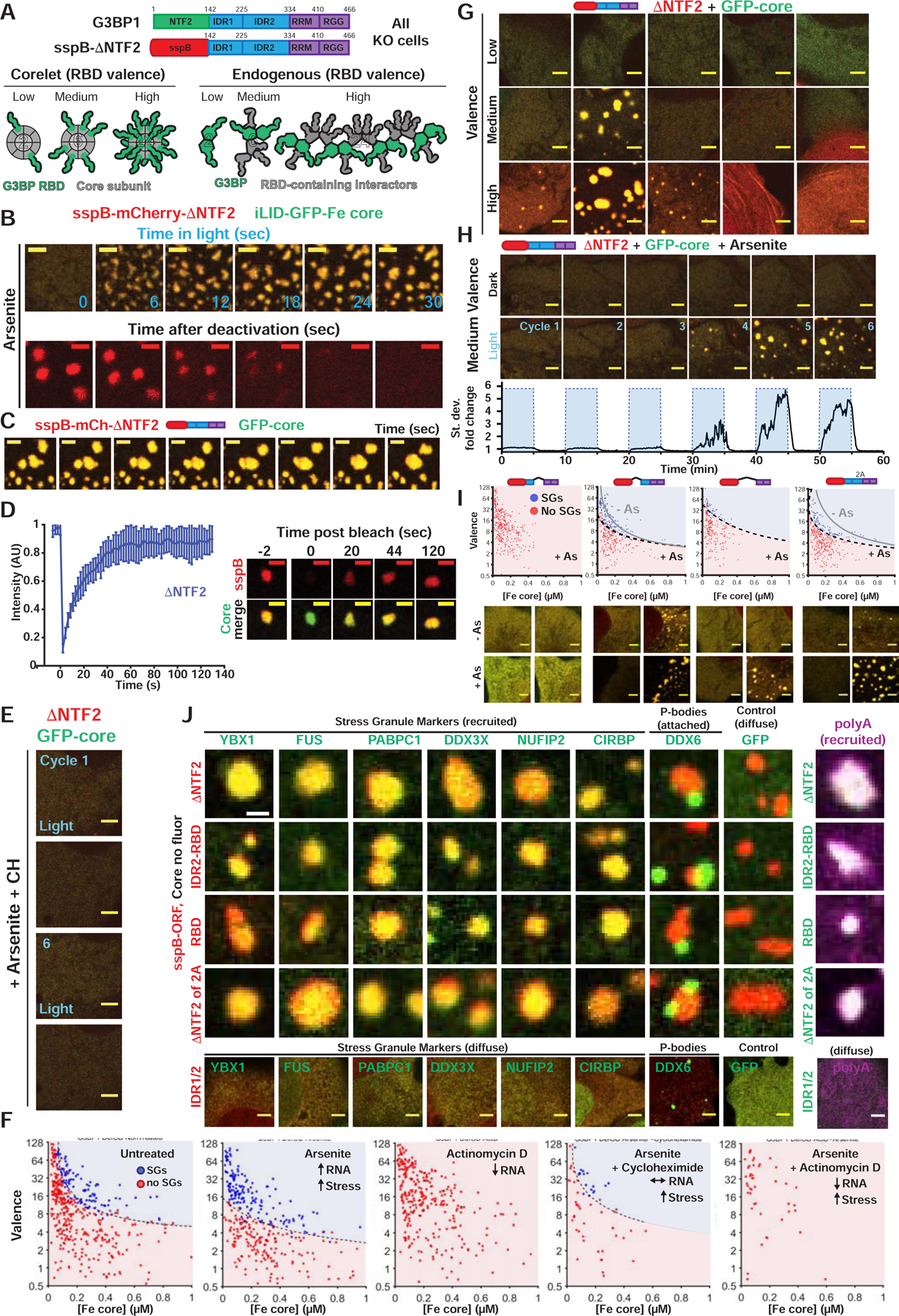
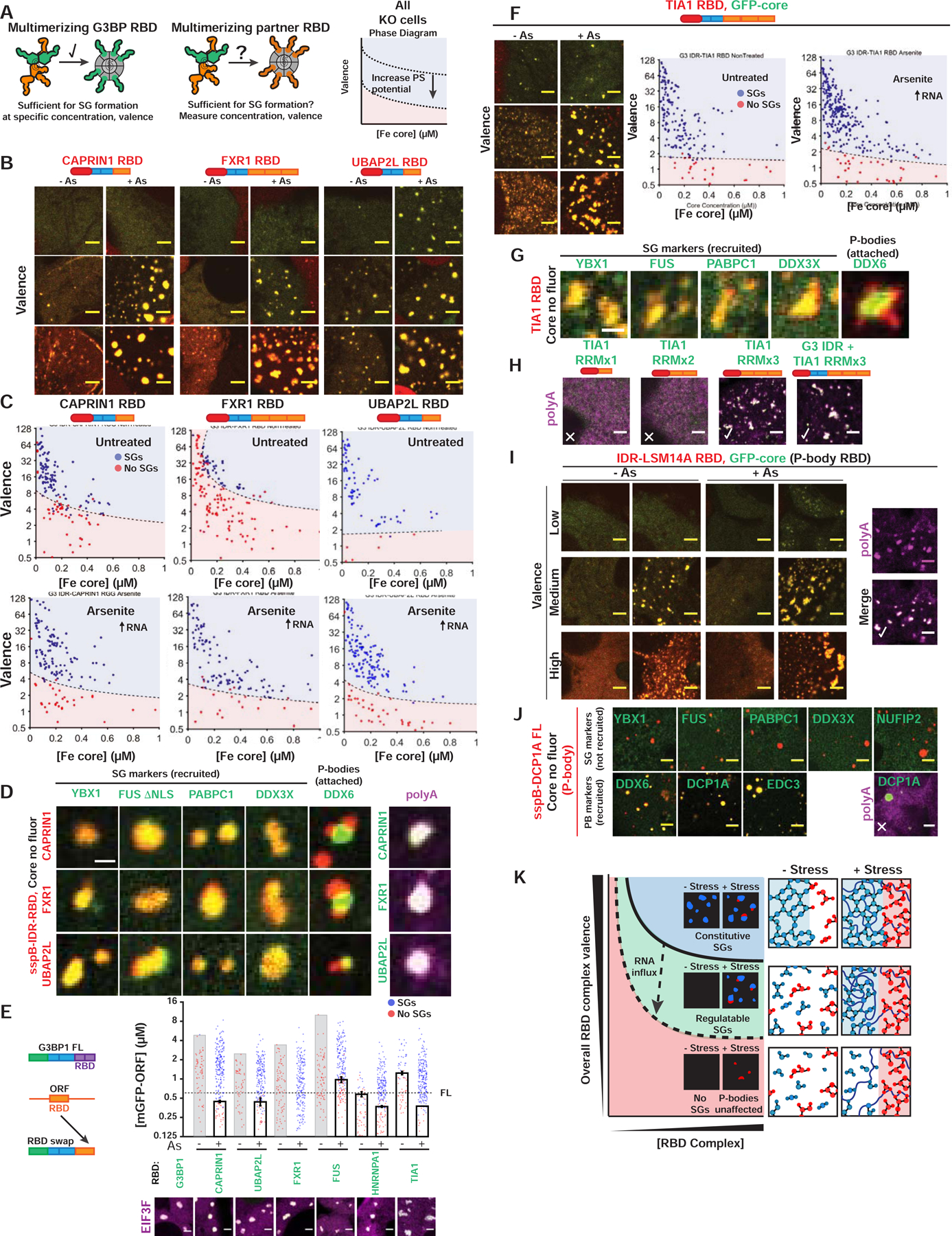
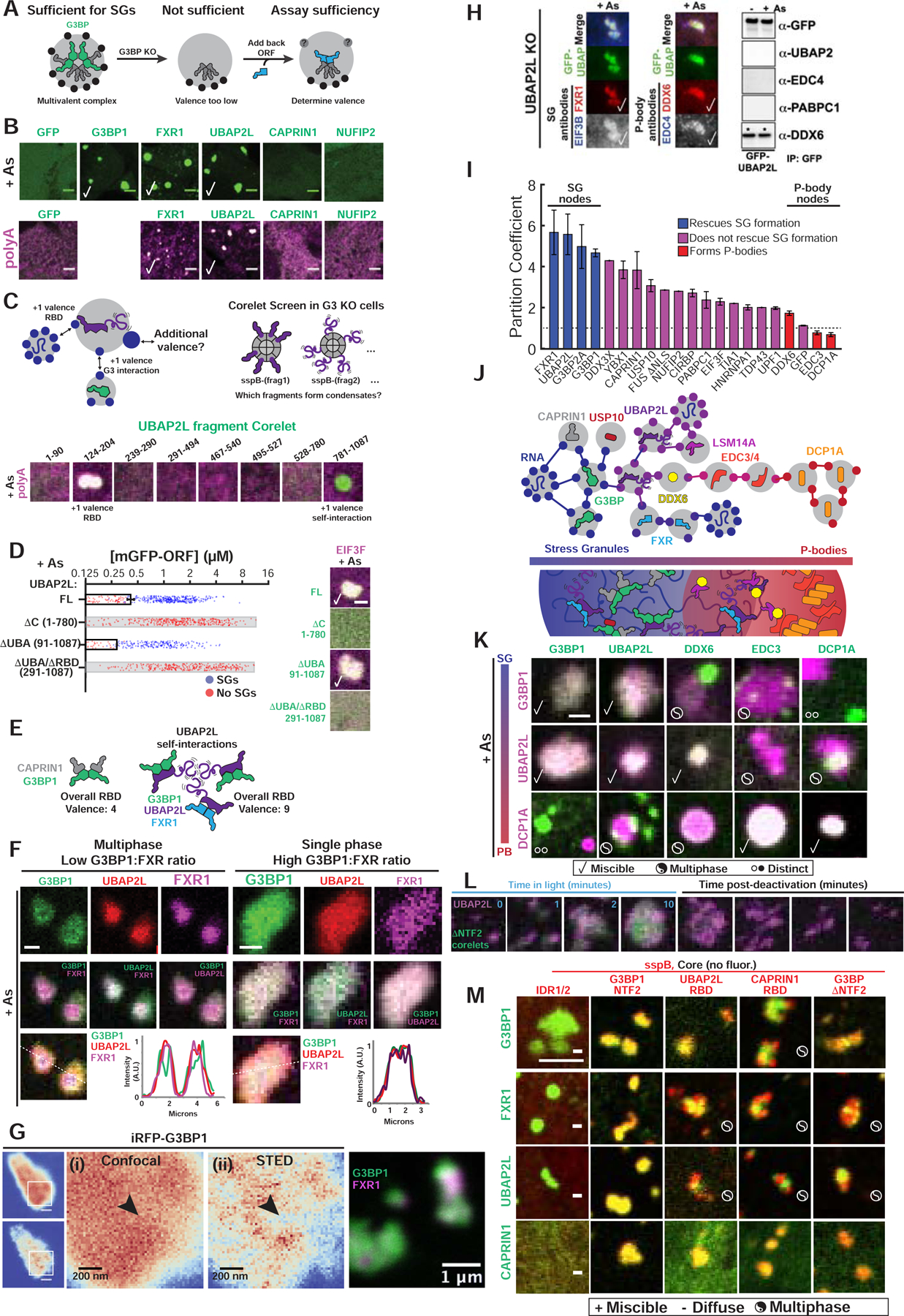
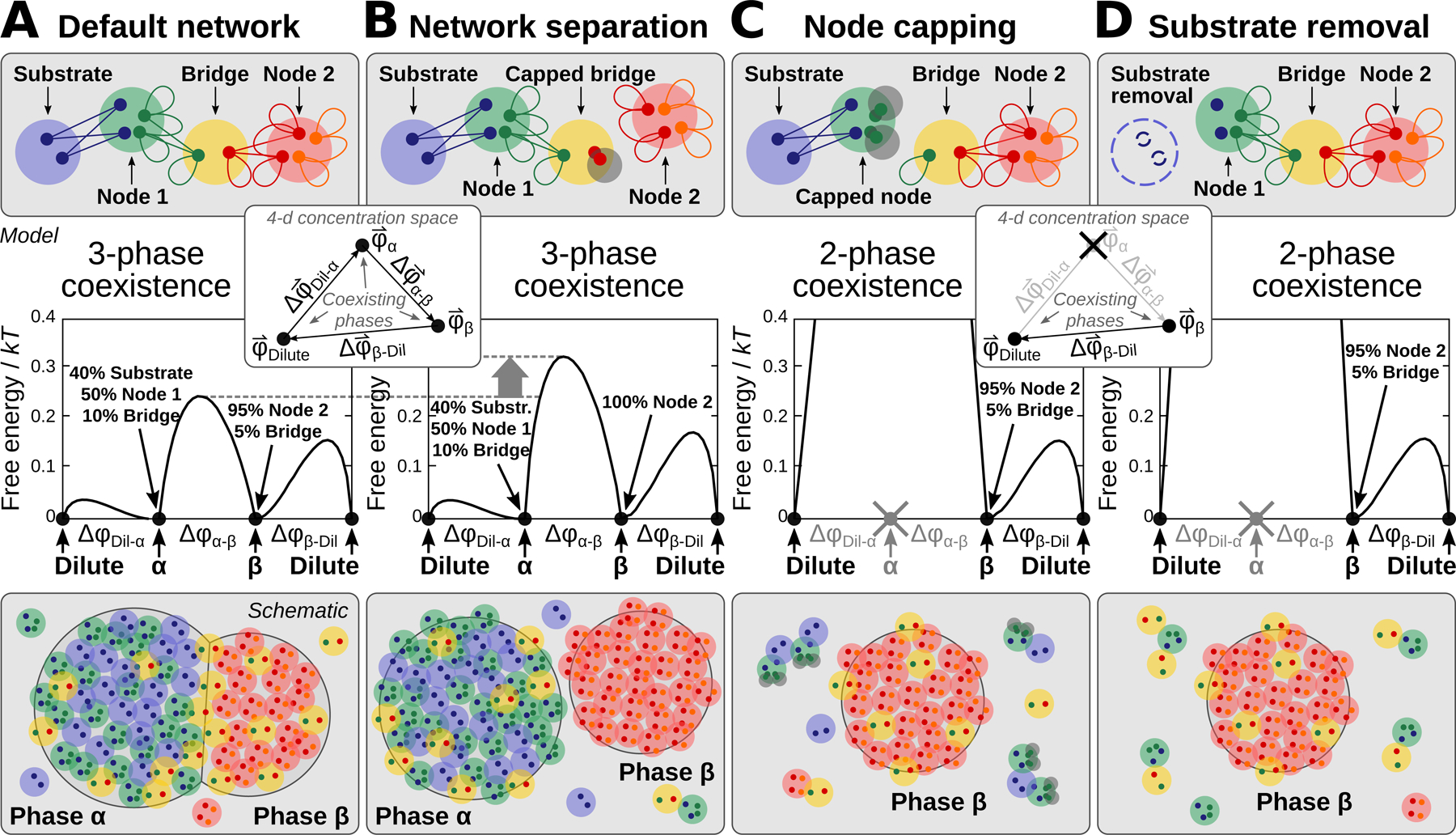
Liquid-liquid phase separation (LLPS) mediates formation of membraneless condensates such as those associated with RNA processing, but the rules that dictate their assembly, substructure, and coexistence with other liquid-like compartments remain elusive. Here, we address the biophysical mechanism of this multiphase organization using quantitative reconstitution of cytoplasmic stress granules (SGs) with attached P-bodies in human cells. Protein-interaction networks can be viewed as interconnected complexes (nodes) of RNA-binding domains (RBDs), whose integrated RNA-binding capacity determines whether LLPS occurs upon RNA influx. Surprisingly, both RBD-RNA specificity and disordered segments of key proteins are non-essential, but modulate multiphase condensation. Instead, stoichiometry-dependent competition between protein networks for connecting nodes determines SG and P-body composition and miscibility, while competitive binding of unconnected proteins disengages networks and prevents LLPS. Inspired by patchy colloid theory, we propose a general framework by which competing networks give rise to compositionally specific and tunable condensates, while relative linkage between nodes underlies multiphase organization.
Keywords: G3BP; P-bodies; RNA binding; UBAP2L; USP10; condensates; membraneless organelles; multiphase; phase separation; stress granules.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests Patent applications have been filed based on this work.
Figures
Comment in
-
Networking and Dynamic Switches in Biological Condensates.Cell. 2020 Apr 16;181(2):228-230. doi: 10.1016/j.cell.2020.03.056. Cell. 2020. PMID: 32302565
References
-
- Adinolfi S, Ramos A, Martin SR, Dal Piaz F, Pucci P, Bardoni B, Mandel JL, and Pastore A (2003). The N-terminus of the fragile X mental retardation protein contains a novel domain involved in dimerization and RNA binding. Biochemistry 42, 10437–10444. - PubMed
-
- Alberti S (2019). Conformational rearrangements within G3BP1 promote the emergence of RNA cross-linking microclusters that drive stress granule formation by condensation. Cell 1–82.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
