

A unique binding mode of Nek2A to the APC/C allows its ubiquitination during prometaphase
- PMID: 32307883
- PMCID: PMC7271329
- DOI: 10.15252/embr.201949831
A unique binding mode of Nek2A to the APC/C allows its ubiquitination during prometaphase
Abstract
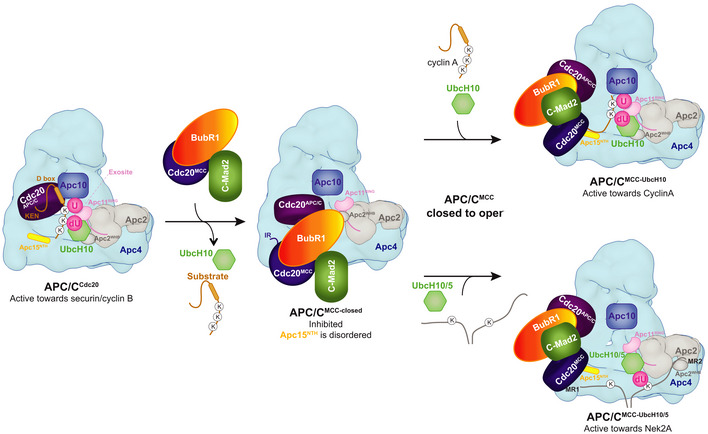
The anaphase-promoting complex (APC/C) is the key E3 ubiquitin ligase which directs mitotic progression and exit by catalysing the sequential ubiquitination of specific substrates. The activity of the APC/C in mitosis is restrained by the spindle assembly checkpoint (SAC), which coordinates chromosome segregation with the assembly of the mitotic spindle. The SAC effector is the mitotic checkpoint complex (MCC), which binds and inhibits the APC/C. It is incompletely understood how the APC/C switches substrate specificity in a cell cycle-specific manner. For instance, it is unclear how in prometaphase, when APC/C activity towards cyclin B and securin is repressed by the MCC, the kinase Nek2A is ubiquitinated. Here, we combine biochemical and structural analysis with functional studies in cells to show that Nek2A is a conformational-specific binder of the APC/C-MCC complex (APC/CMCC ) and that, in contrast to cyclin A, Nek2A can be ubiquitinated efficiently by the APC/C in conjunction with both the E2 enzymes UbcH10 and UbcH5. We propose that these special features of Nek2A allow its prometaphase-specific ubiquitination.
Keywords: E3 ligase; anaphase-promoting complex; cell cycle; cryo-EM; spindle assembly checkpoint.
© 2020 The Authors.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
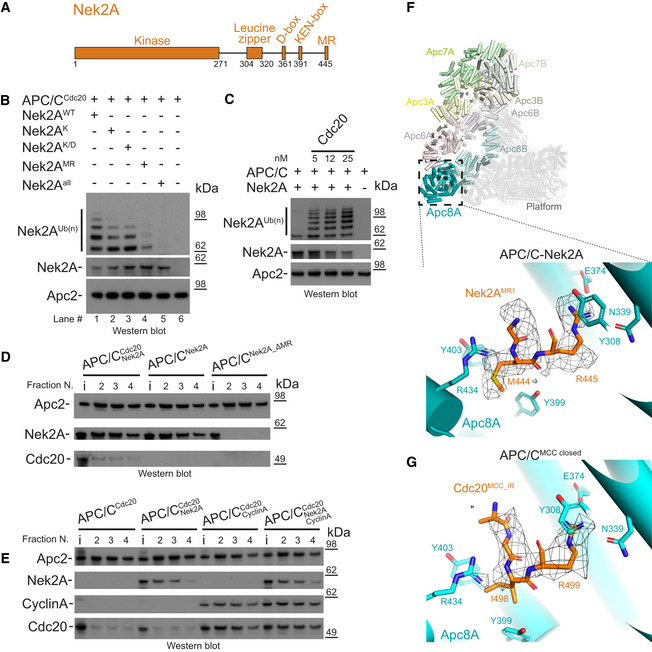
Schematic representation of the domain and degron architecture of Nek2A.
The MR tail is the main contributor for Nek2A ubiquitination. Ubiquitination reactions performed with APC/CCdc20 for Nek2A wild‐type (Nek2AWT) and degron mutants. Nek2AK is the KEN‐box mutant (391KEN393/KAA), Nek2AK/D is the (KEN‐ and D‐box mutant (391KEN393/KAA, 361RKFL364/AKFA), and Nek2AMR is the 443ΔMR mutant.
The APC/C requires the Cdc20 coactivator subunit to ubiquitinate Nek2A. Nek2A ubiquitination reactions performed with the APC/C in the absence and presence of increasing concentrations of Cdc20 using the E2 UbcH10.
The MR tail is essential for Nek2A binding to the APC/C, whereas in contrast Cdc20 is dispensable. Size‐exclusion chromatography peak fractions of APC/C complexes with Nek2A. The input material (13% of total) is indicated (i). Peak fractions are numbered in respect of the chromatograms in Fig EV1.
Nek2A binding does not stabilize binding of the coactivator subunit Cdc20 to the APC/C and does not compete with cyclin A for binding to the APC/C. Size‐exclusion chromatography peak fractions of APC/C complexes with Nek2A.
Top: Cylinder representation of the APC/C–Nek2A complex structure. The TPR lobe subunits are highlighted: Apc8 (cyan), Apc6 (pink), Apc3 (yellow) and Apc7 (green). The platform subunits and the small subunits from the TPR lobe are shown in transparency. Bottom: cryo‐EM density of the Nek2AMR tail dipeptide and details of the Nek2AMR tail (orange) binding site on Apc8A.
Details of the IR tail of Cdc20MCC (orange) bound to Apc8A in APC/CMCC‐closed. This is the same site as the Nek2AMR tail binding site shown above. Shown is the cryo‐EM density for the Cdc20MCC IR tail and contacting residues of Apc8A.
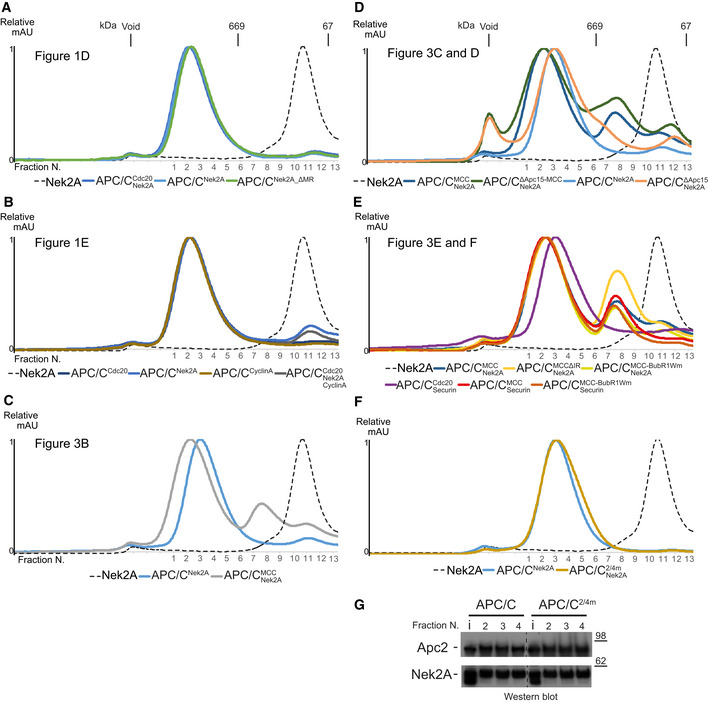
- A–E
Size‐exclusion chromatograms of APC/C complexes in this study. The corresponding peak fractions SDS–PAGE gels are shown in the main figures.
- F, G
Size‐exclusion chromatography chromatograms (F) and SDS–PAGE gels (G) of either APC/C wild‐type or 2/4 m complexes with Nek2A.
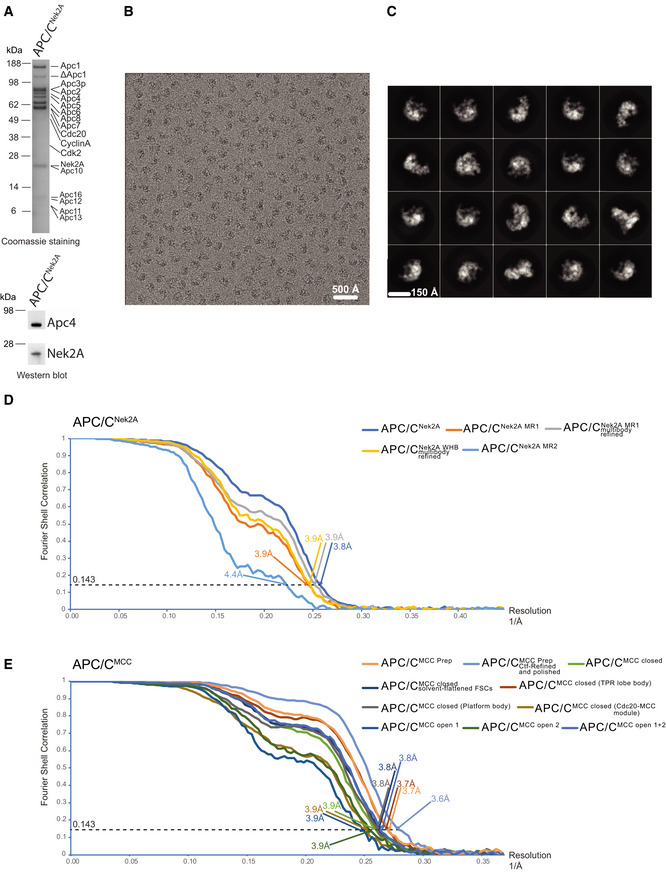
SDS–PAGE gels (stained with Coomassie blue) of the gel filtration peak fraction from the APC/C–Nek2A complex preparation used in this study. Western blot against Nek2A confirms the presence of the Nek2A 301–406 construct.
A typical cryo‐EM micrograph of APC/C–Nek2A representative of 6,448 micrographs. Scale bar: 500 Å.
Gallery of two‐dimensional class averages of APC/CNek2A showing different views representative of 50 two‐dimensional classes. Scale bar: 150 Å.
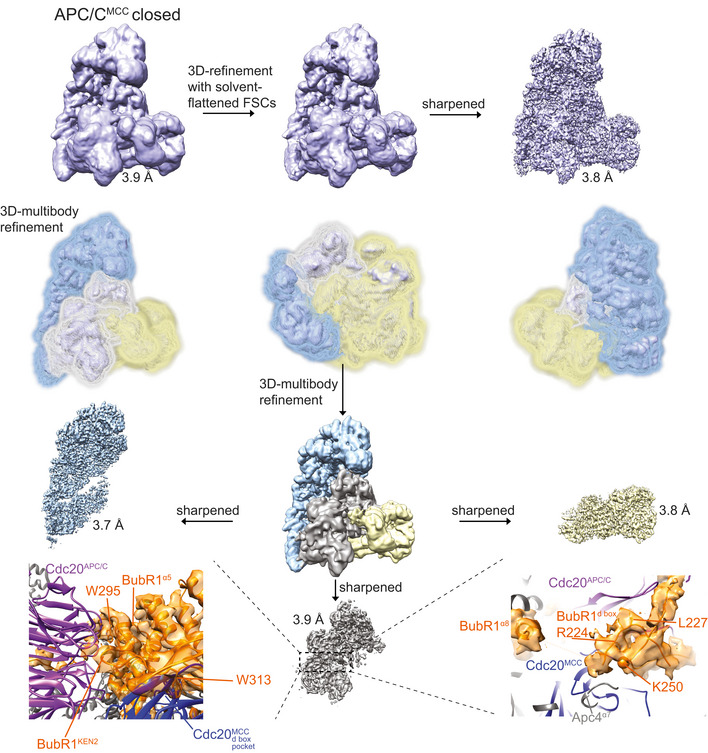
Fourier shell correlation (FSC) curves are shown for all the cryo‐EM reconstructions of APC/CNek2A determined in this study.
Fourier shell correlation (FSC) curves are shown for all the cryo‐EM reconstructions of APC/CMCC determined in this study using cryo‐EM data as published 14.
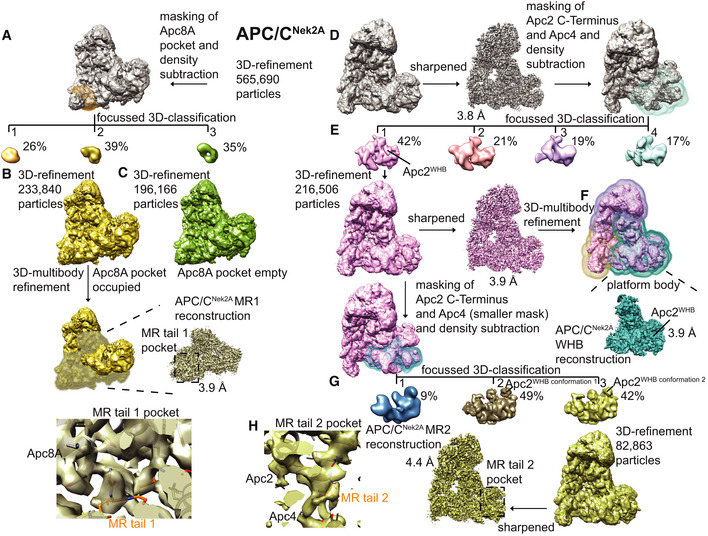
- A–C
Workflow for focussed 3D classification and multi‐body refinement for 3D reconstruction and refinement of the cryo‐EM density for the MR tail 1 binding site on Apc8A.
- D–H
Workflow for focussed 3D classification and multi‐body refinement for 3D reconstruction and refinement of the cryo‐EM density for the repositioned APC2WHB domain and for the MR tail 2 binding site on Apc2‐Apc4. Classes 2 and 3 shown in (G) differed in the orientation of the APC2WHB domain, and the occupancy of the MR tail 2 was highest in class 3.
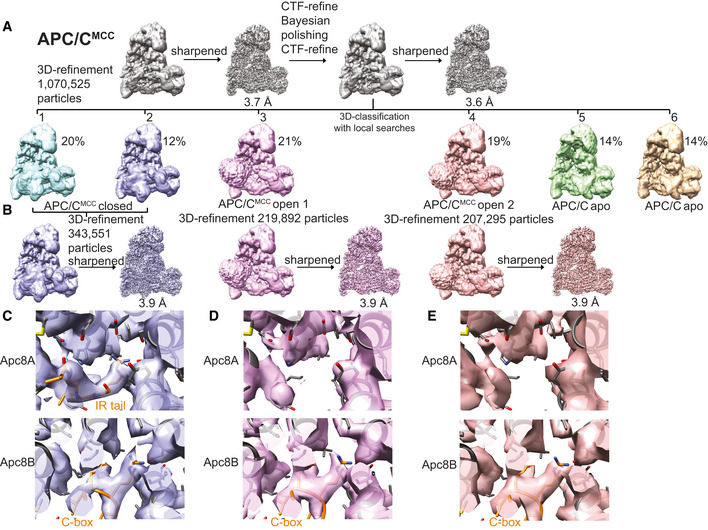
- A
3D class averages obtained by classification using local searches (see Materials and Methods) are shown for obtaining the APC/CMCC reconstructions.
- B
Particles from classes 1 and 2 were refined together to obtain the APC/CMCC closed reconstruction. Particles from classes 3 and 4 were separately refined to obtain the APC/CMCC open 1 and 2 reconstructions, respectively. These latter classes were slightly different in the orientation of the Cdc20‐MCC module.
- C
Details of the IR tail and C‐box binding site of the APC/CMCC‐closed reconstruction.
- D, E
(D). Details of the IR tail and C‐box binding site of the APC/CMCC‐open−1 reconstruction and (E) of the APC/CMCC‐open−2 reconstruction. Neither of the two APC/CMCC‐open reconstructions shows clear density for the Cdc20MCC IR tail.
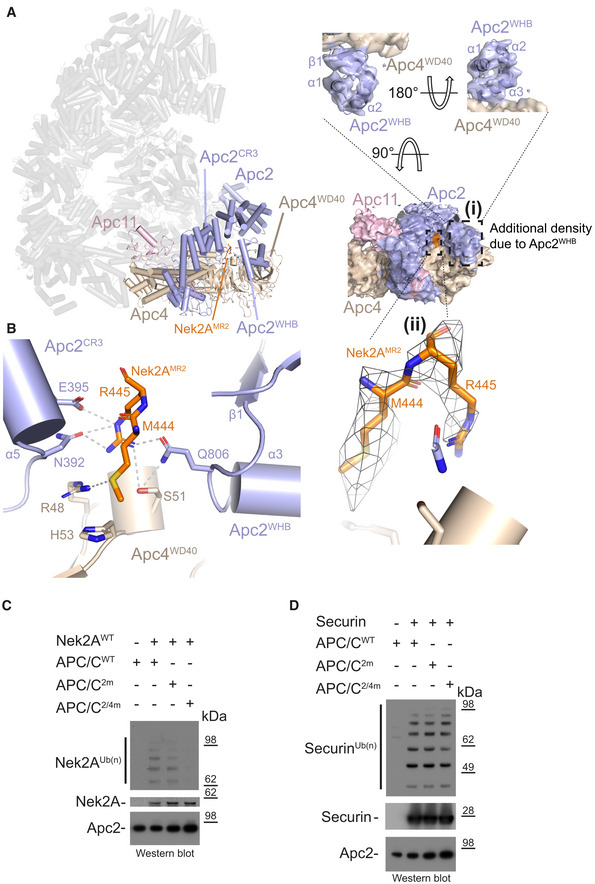
Left: Cylinder representation of the APC/C–Nek2A complex structure. Apc2 (light blue) and Apc4 (light brown) are highlighted, and the rest of the APC/C is shown in transparency. (i) Two different views of cryo‐EM density for the Apc2 and Apc4 subunits. Secondary structure elements of the Apc2WHB domain are highlighted. (ii) Cryo‐EM density for the second MR tail of Nek2A.
Details of the second MR tail binding site formed by the Apc2 (including the repositioned Apc2WHB) and Apc4 subunits. Nek2AMR2 is shown in orange.
Nek2A ubiquitination reactions performed with either APC/CCdc20 wild type (APC/CWT) or mutants. APC/C2m is the N392A/E395A mutant of Apc2. APC/C2/4m is the N392A/E395A/R48A/H53A/S51A mutant of Apc2 and Apc4.
Securin ubiquitination reactions performed with either APC/CCdc20 wild type (APC/CWT) or mutants.
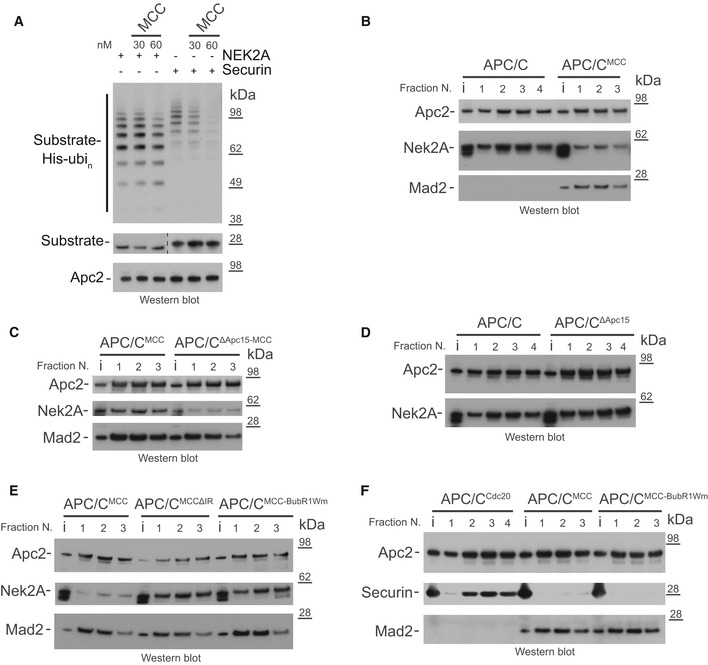
- A
Ubiquitination reactions of either Nek2A or securin by the APC/CCdc20 and the effect of increasing concentrations of the MCC.
- B–F
Size‐exclusion chromatography peak fractions of APC/C complexes with either Nek2A (B‐E) or securin (F). APC/CΔApc15 is the Apc15 deleted mutant APC/C, and MCCΔIR is the Cdc20 IR tail deleted mutant MCC. MCC‐BubR1Wm is the MCC mutant at the APC2WHB binding surface on BubR1 14. Chromatograms are shown in Fig EV1.
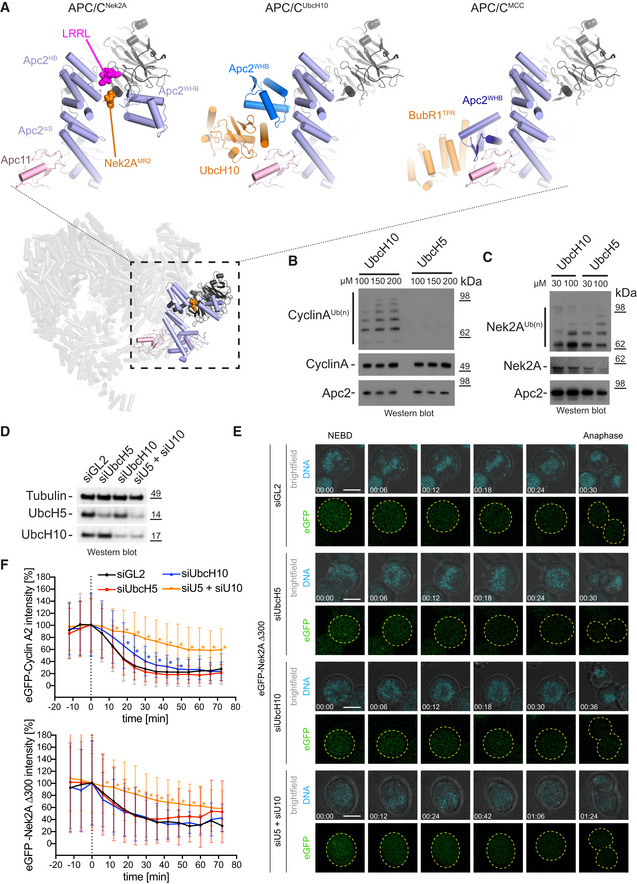
Bottom: Overview of the APC/C–Nek2A structure is shown coloured as in Fig 2. Top: Close up of the Apc2 and Apc4 binding site for Nek2A in the APC/C–Nek2A structure (this study, left), APC/CUbcH10‐Hsl1 structure 27 and APC/CMCC structure 14.
Ubiquitination reactions of cyclin A by the APC/CCdc20 in the presence of increasing concentrations of either UbcH10 or UbcH5.
Ubiquitination reactions of Nek2A by the APC/CCdc20 in the presence of increasing concentrations of either UbcH10 or UbcH5.
Western blot of Hek293 cells showing the siRNA‐mediated depletion of UbcH5, UbcH10 or both E2 enzymes together. Tubulin serves as a loading control.
Exemplary still images from time courses between NEBD and anaphase of eGFP‐Nek2A degradation in HEK cells. Cells were either treated with siGL2 as control or depleted of the indicated E2 enzymes. The chromosomes are coloured in cyan and eGFP‐Nek2A in green, with the outline of the cells are indicated with dashed yellow lines. Time is given as hh:mm. Scale bar 10 μm. See also [Link], [Link], [Link], [Link] and Appendix Fig S1.
Degradation profiles of eGFP‐cyclin A2 (top) and eGFP‐Nek2A (bottom) in HEK cells during mitosis. The time point of NEBD is marked at 0 min in the graphs. Asterisks indicate values that are significantly different from the same time point of the siGL2 control as determined by a Mann–Whitney U‐test (the statistics are listed in Table EV2). Mean ± SD is shown. The number of cells analysed are N = 66 (Cyclin A2 siGL2), 52 (Cyclin A2 siUbcH5), 28 (Cyclin A2 siUbcH10), 27 (Cyclin A2 siU5/siU10), 80 (Nek2A siGL2), 64 (Nek2A siUbcH5), 21 (Nek2A siUbcH10) and 32 (Nek2A siU5/siU10). All data are from at least two biological replicates. See also [Link], [Link], [Link], [Link].
Comment in
-
Escape from the checkpoint: Nek2A binds a unique conformation of the APC/C-MCC complex.EMBO Rep. 2020 Jun 4;21(6):e50494. doi: 10.15252/embr.202050494. Epub 2020 May 19. EMBO Rep. 2020. PMID: 32431068 Free PMC article.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
