

Disentangling the mechanisms shaping the surface ocean microbiota
- PMID: 32312331
- PMCID: PMC7171866
- DOI: 10.1186/s40168-020-00827-8
Disentangling the mechanisms shaping the surface ocean microbiota
Abstract
Background: The ocean microbiota modulates global biogeochemical cycles and changes in its configuration may have large-scale consequences. Yet, the underlying ecological mechanisms structuring it are unclear. Here, we investigate how fundamental ecological mechanisms (selection, dispersal and ecological drift) shape the smallest members of the tropical and subtropical surface-ocean microbiota: prokaryotes and minute eukaryotes (picoeukaryotes). Furthermore, we investigate the agents exerting abiotic selection on this assemblage as well as the spatial patterns emerging from the action of ecological mechanisms. To explore this, we analysed the composition of surface-ocean prokaryotic and picoeukaryotic communities using DNA-sequence data (16S- and 18S-rRNA genes) collected during the circumglobal expeditions Malaspina-2010 and TARA-Oceans.
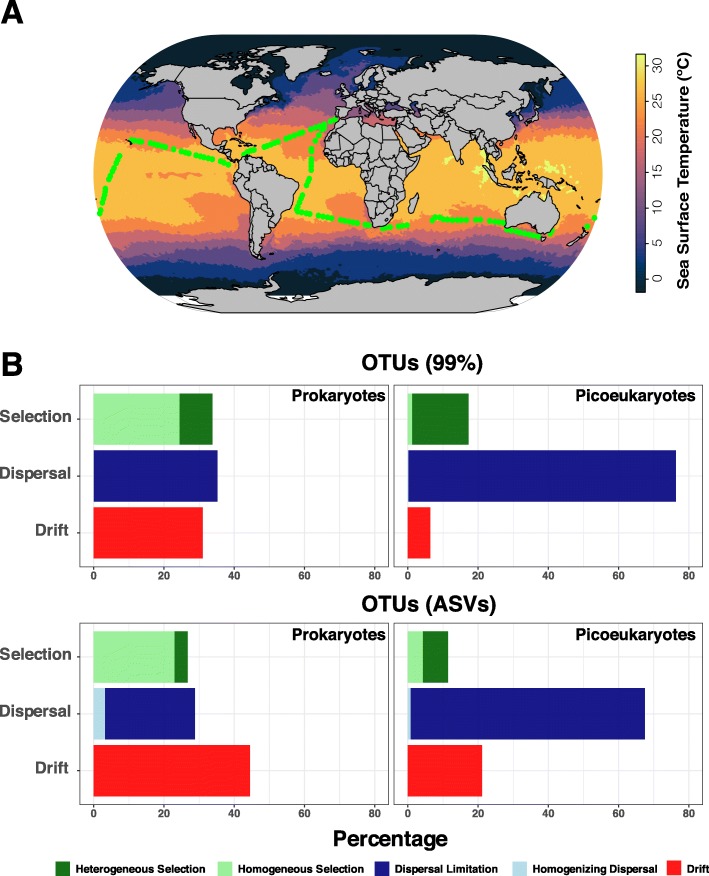
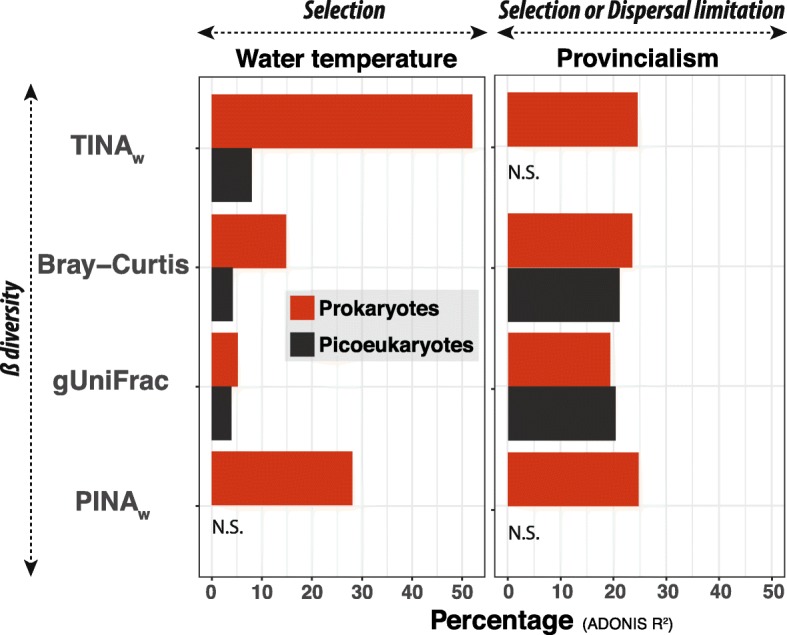
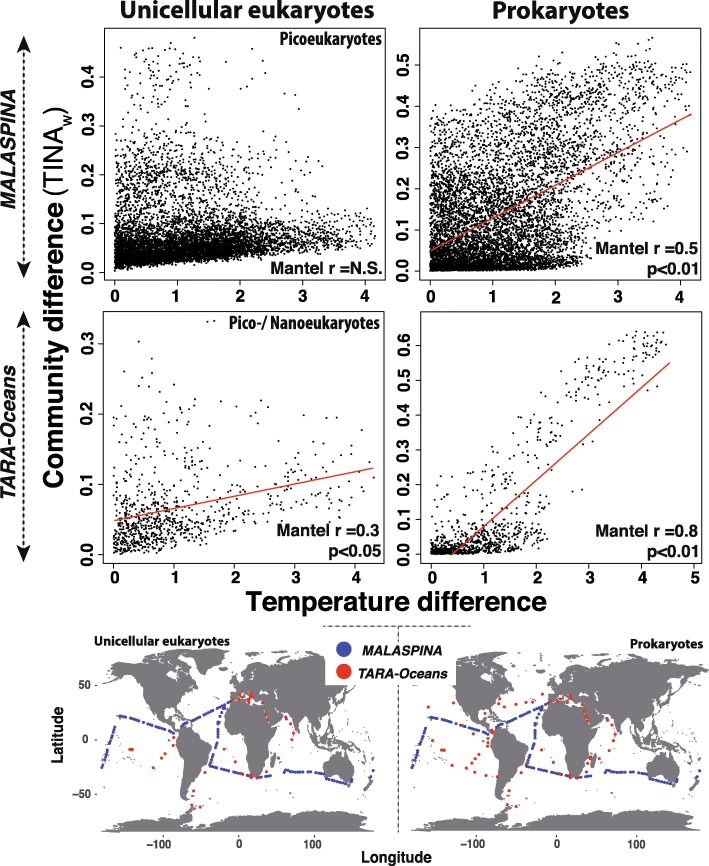
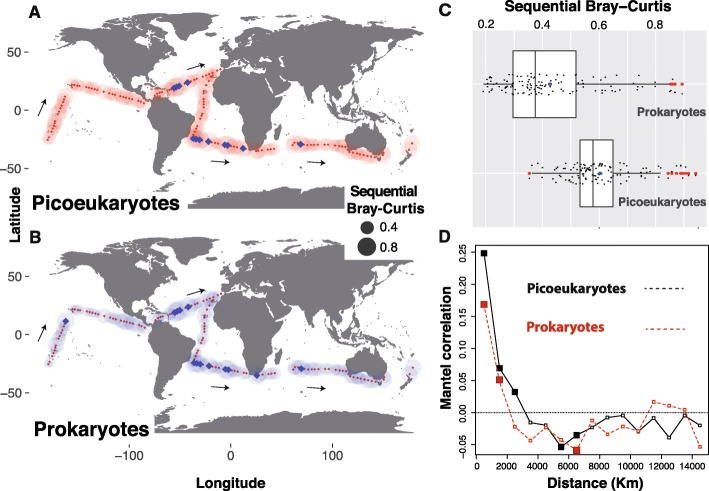
Results: We found that the two main components of the tropical and subtropical surface-ocean microbiota, prokaryotes and picoeukaryotes, appear to be structured by different ecological mechanisms. Picoeukaryotic communities were predominantly structured by dispersal-limitation, while prokaryotic counterparts appeared to be shaped by the combined action of dispersal-limitation, selection and drift. Temperature-driven selection appeared as a major factor, out of a few selected factors, influencing species co-occurrence networks in prokaryotes but not in picoeukaryotes, indicating that association patterns may contribute to understand ocean microbiota structure and response to selection. Other measured abiotic variables seemed to have limited selective effects on community structure in the tropical and subtropical ocean. Picoeukaryotes displayed a higher spatial differentiation between communities and a higher distance decay when compared to prokaryotes, consistent with a scenario of higher dispersal limitation in the former after considering environmental heterogeneity. Lastly, random dynamics or drift seemed to have a more important role in structuring prokaryotic communities than picoeukaryotic counterparts.
Conclusions: The differential action of ecological mechanisms seems to cause contrasting biogeography, in the tropical and subtropical ocean, among the smallest surface plankton, prokaryotes and picoeukaryotes. This suggests that the idiosyncrasy of the main constituents of the ocean microbiota should be considered in order to understand its current and future configuration, which is especially relevant in a context of global change, where the reaction of surface ocean plankton to temperature increase is still unclear. Video Abstract.
Keywords: Community structure; Dispersal; Drift; Ecological processes; Microbiota; Ocean; Picoeukaryotes; Plankton; Prokaryotes; Selection.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Li WKW. Primary production of prochlorophytes, cyanobacteria, and eucaryotic ultraphytoplankton: Measurements from flow cytometric sorting. Limnol Oceanography. 1994;39(1):169–175. doi: 10.4319/lo.1994.39.1.0169. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
