

The regulation and functions of DNA and RNA G-quadruplexes
- PMID: 32313204
- PMCID: PMC7115845
- DOI: 10.1038/s41580-020-0236-x
The regulation and functions of DNA and RNA G-quadruplexes
Abstract
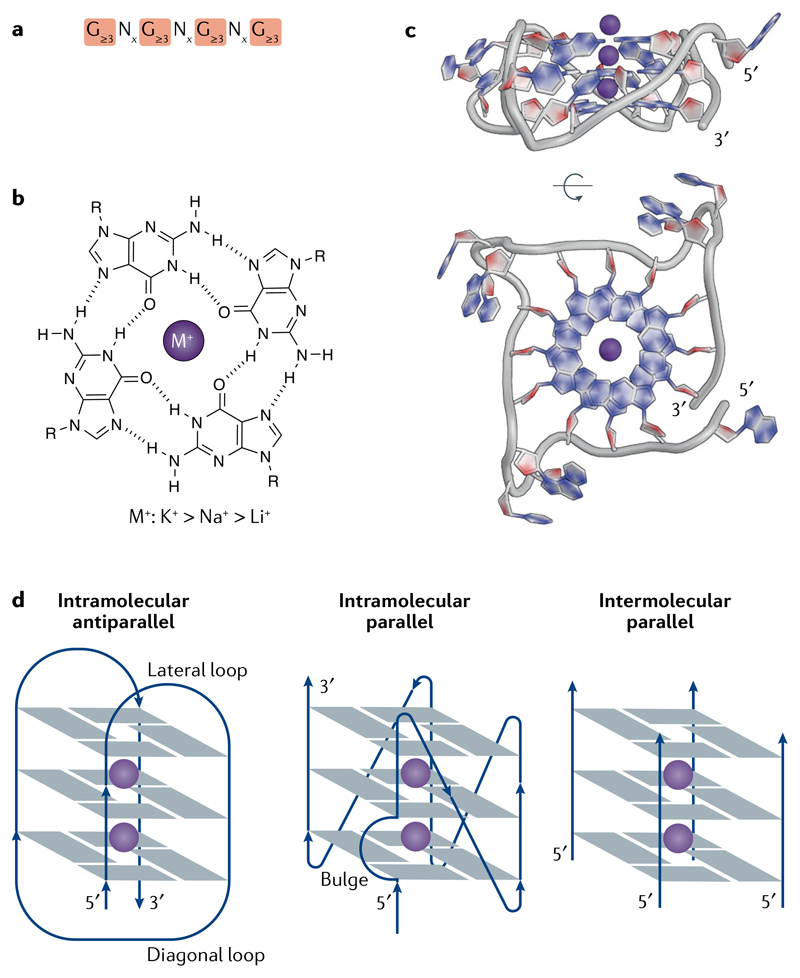
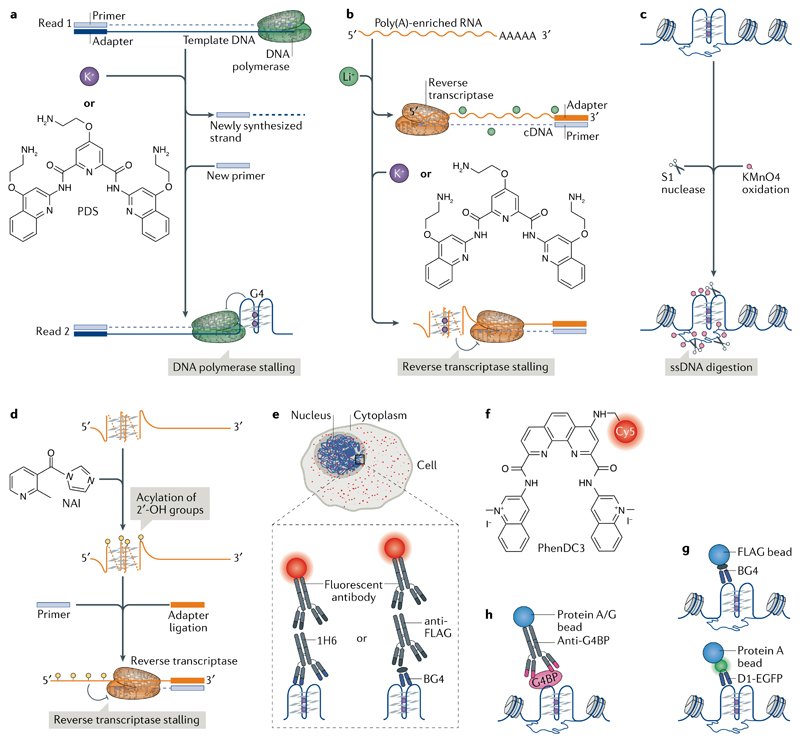
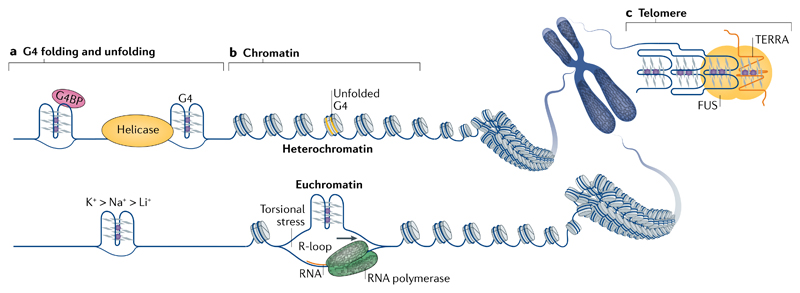
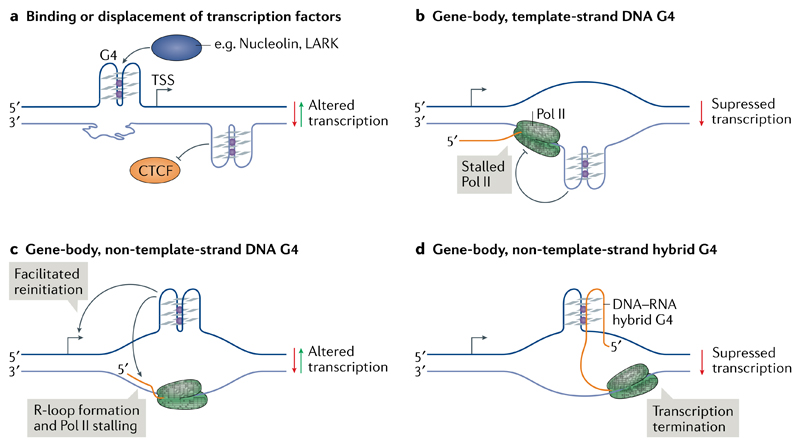
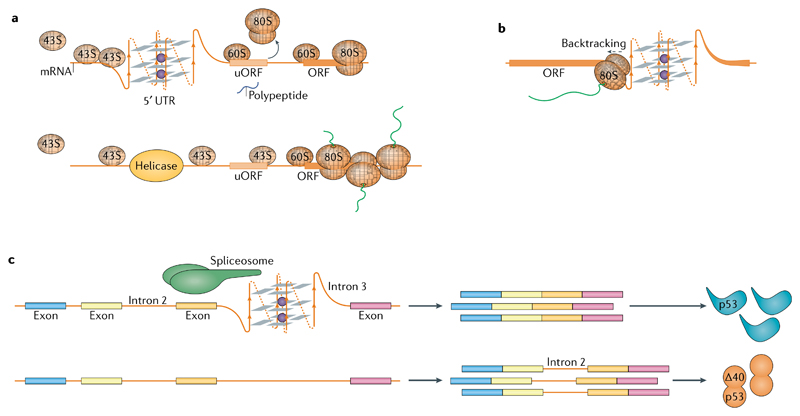
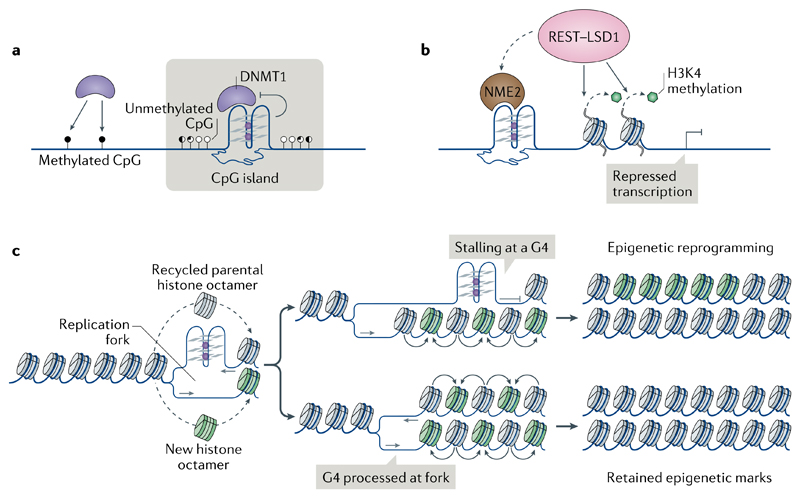
DNA and RNA can adopt various secondary structures. Four-stranded G-quadruplex (G4) structures form through self-recognition of guanines into stacked tetrads, and considerable biophysical and structural evidence exists for G4 formation in vitro. Computational studies and sequencing methods have revealed the prevalence of G4 sequence motifs at gene regulatory regions in various genomes, including in humans. Experiments using chemical, molecular and cell biology methods have demonstrated that G4s exist in chromatin DNA and in RNA, and have linked G4 formation with key biological processes ranging from transcription and translation to genome instability and cancer. In this Review, we first discuss the identification of G4s and evidence for their formation in cells using chemical biology, imaging and genomic technologies. We then discuss possible functions of DNA G4s and their interacting proteins, particularly in transcription, telomere biology and genome instability. Roles of RNA G4s in RNA biology, especially in translation, are also discussed. Furthermore, we consider the emerging relationships of G4s with chromatin and with RNA modifications. Finally, we discuss the connection between G4 formation and synthetic lethality in cancer cells, and recent progress towards considering G4s as therapeutic targets in human diseases.
Conflict of interest statement
S.B. is a founder and shareholder of Cambridge Epigenetix Ltd.
Figures
References
-
- Bang I. Untersuchungen über die Guanylsäure. Biochemische. 1910;26:293–311.
-
- Sen D, Gilbert W. Formation of parallel four-stranded complexes by guanine-rich motifs in DNA and its implications for meiosis. Nature. 1988;334:364–366. [ This paper is an early demonstration of a G4 comprising stacked tetrads with interconnecting loop sequences performed using chemical mapping and providing biological insight. ] - PubMed
-
- Kwok CK, Merrick CJ. G-Quadruplexes: prediction, characterization, and biological application. Trends Biotechnol. 2017;35:997–1013. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
