

Selective PP2A Enhancement through Biased Heterotrimer Stabilization
- PMID: 32315618
- PMCID: PMC7243596
- DOI: 10.1016/j.cell.2020.03.038
Selective PP2A Enhancement through Biased Heterotrimer Stabilization
Abstract
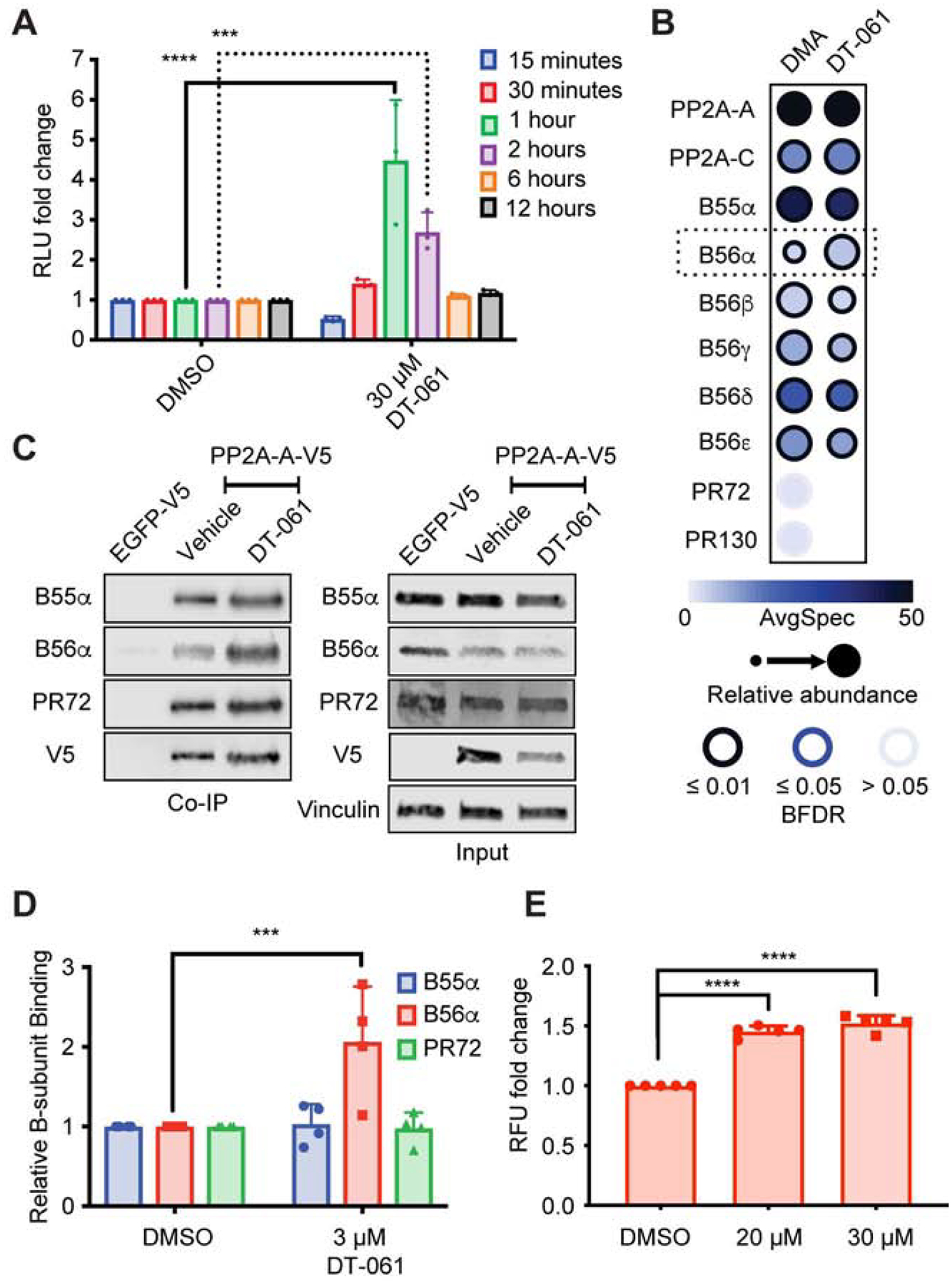
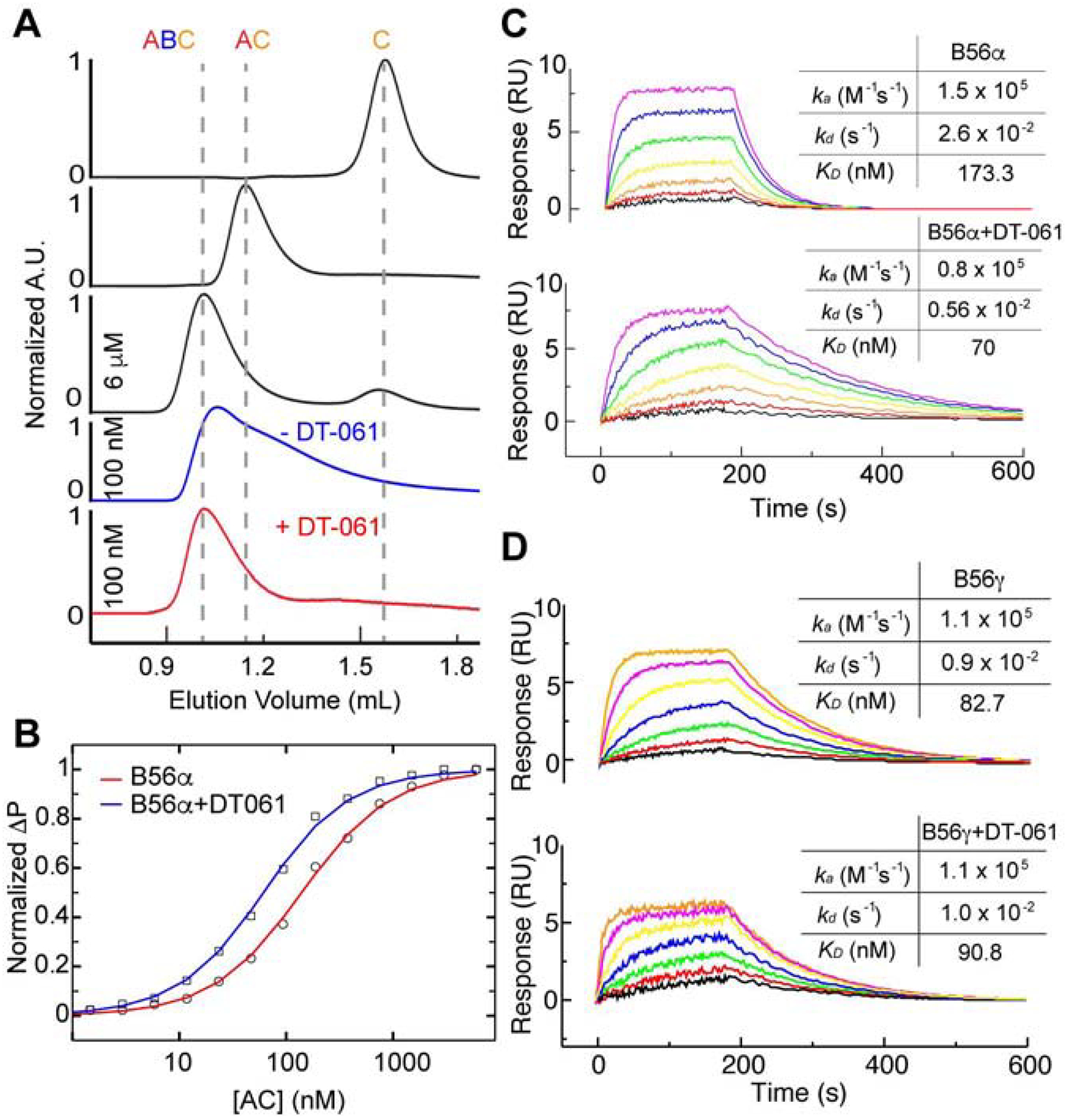
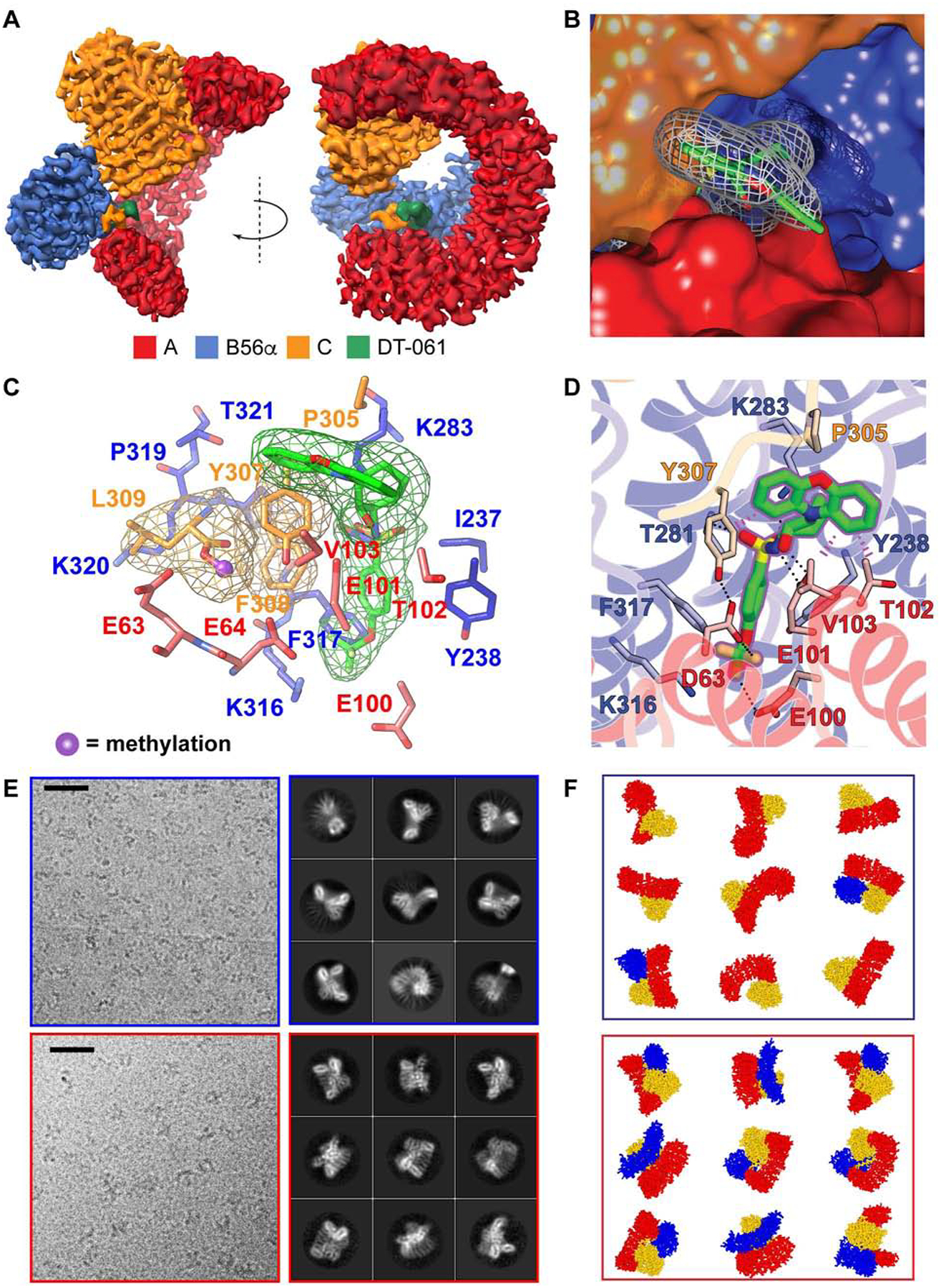
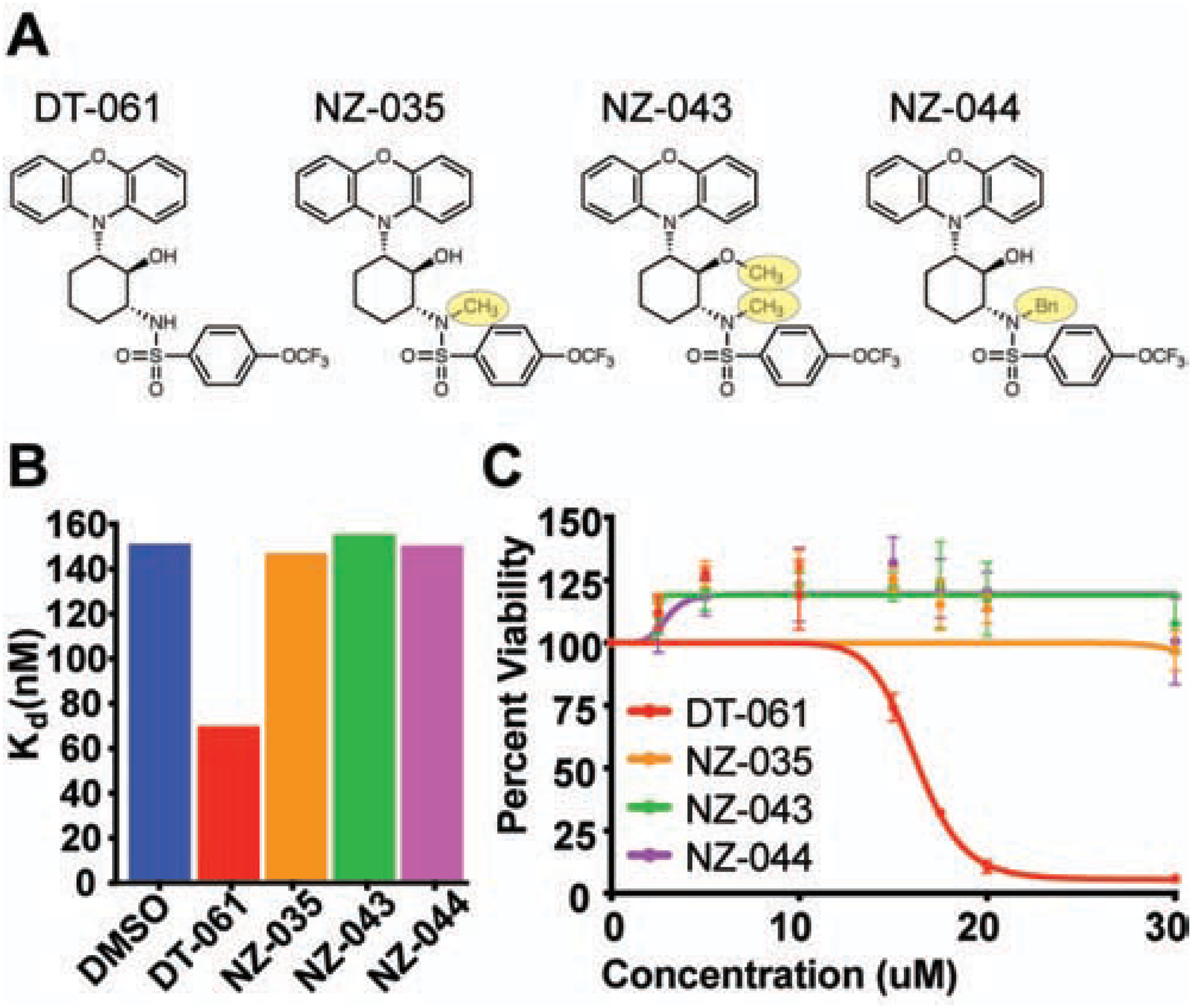
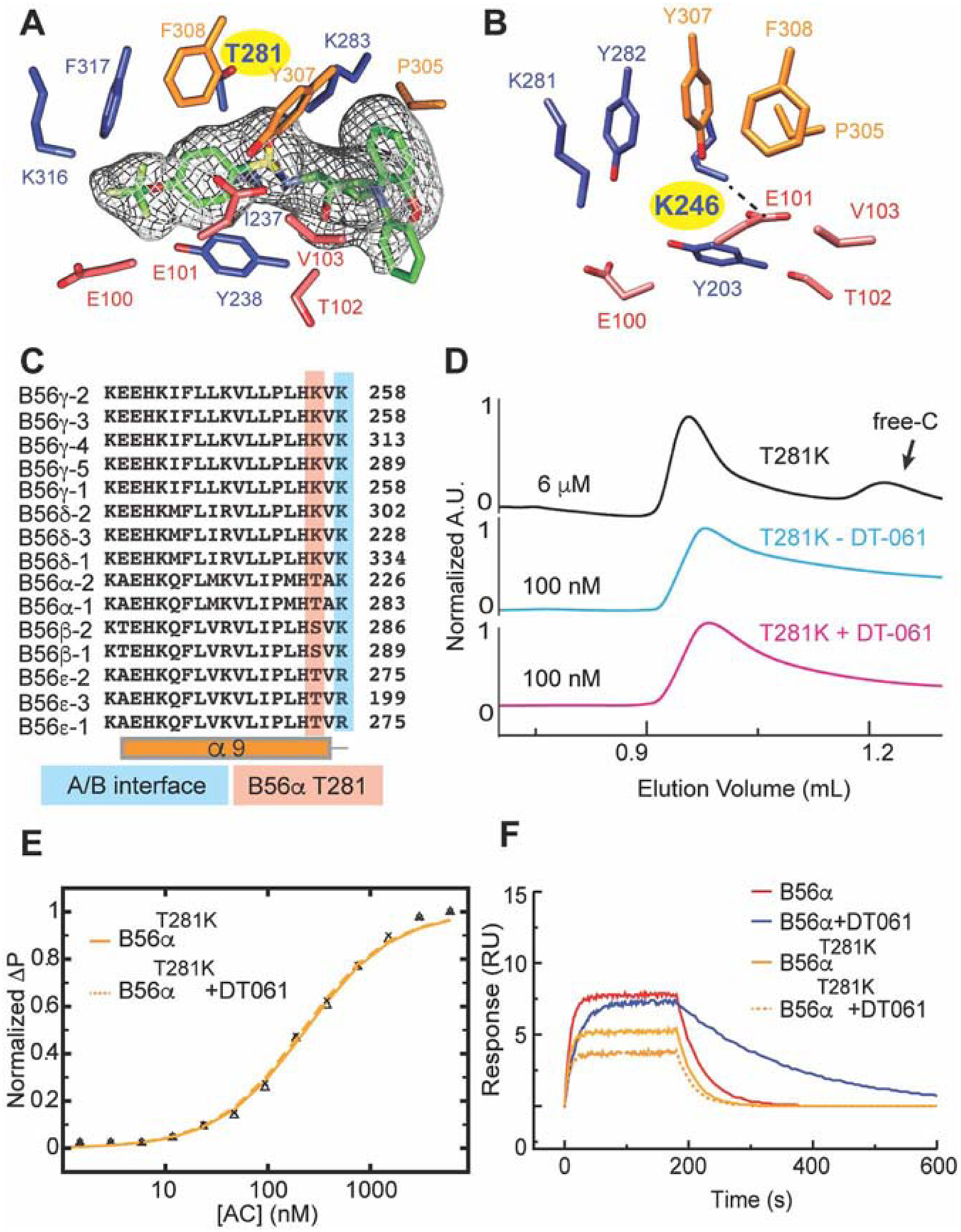
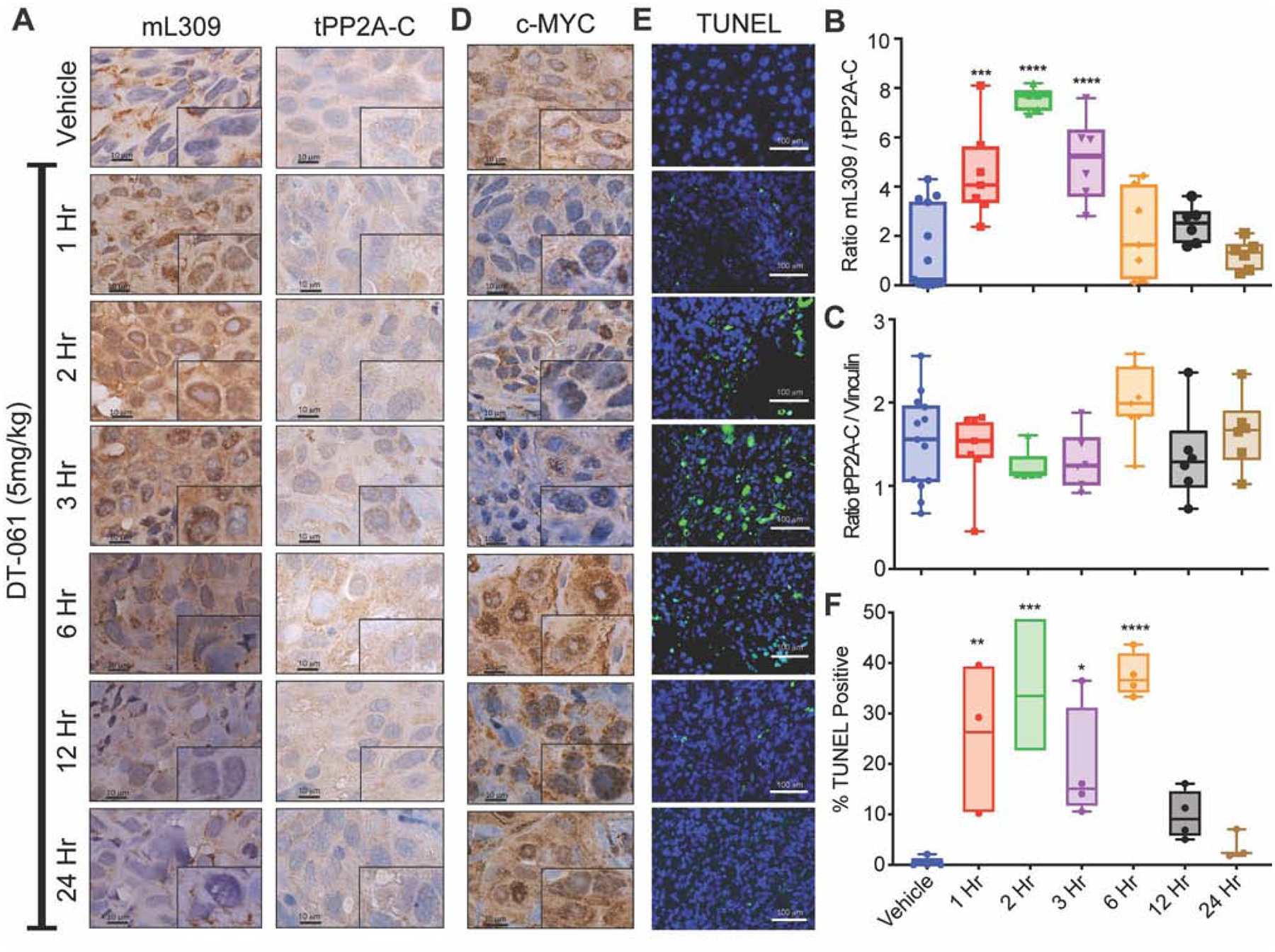
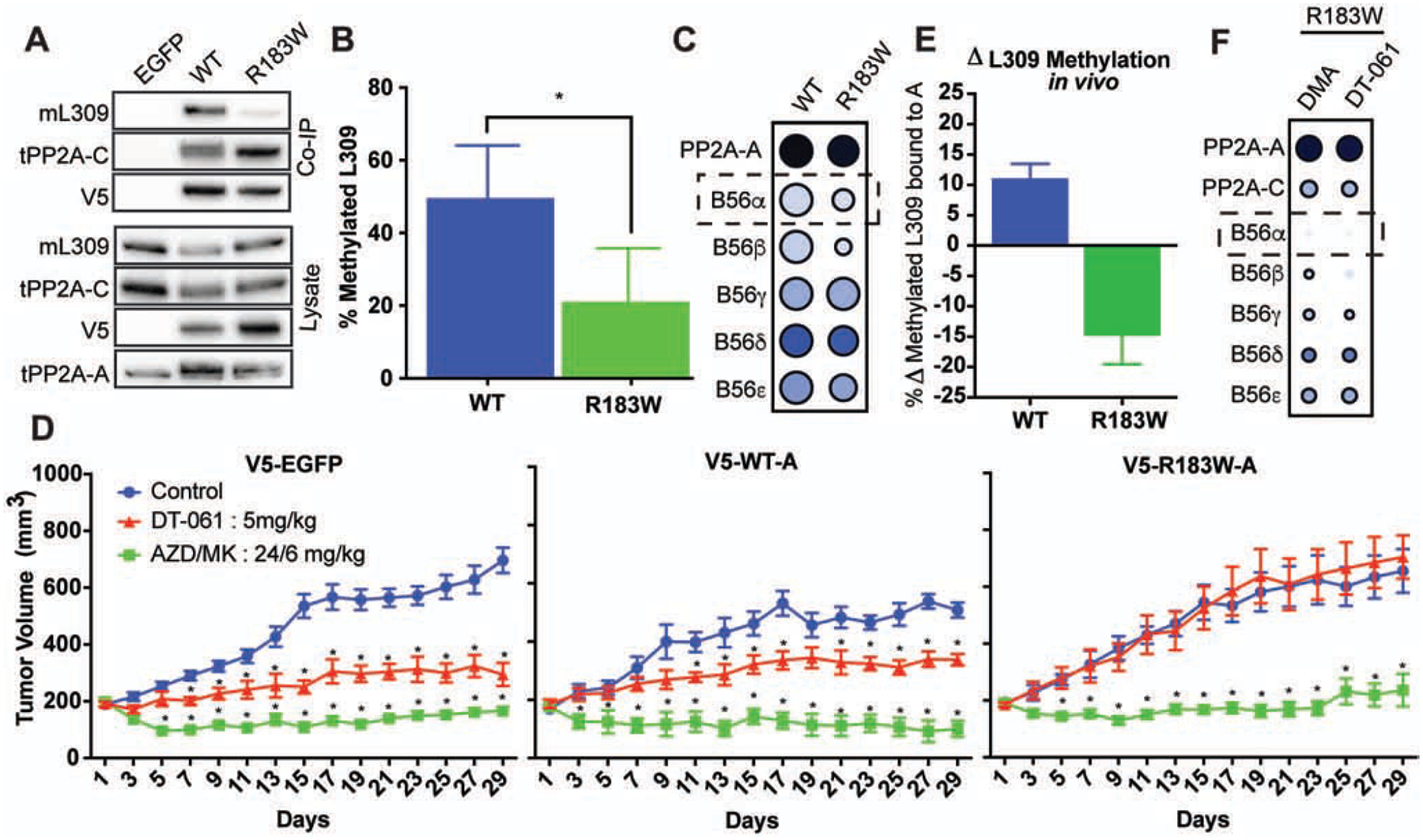
Impairment of protein phosphatases, including the family of serine/threonine phosphatases designated PP2A, is essential for the pathogenesis of many diseases, including cancer. The ability of PP2A to dephosphorylate hundreds of proteins is regulated by over 40 specificity-determining regulatory "B" subunits that compete for assembly and activation of heterogeneous PP2A heterotrimers. Here, we reveal how a small molecule, DT-061, specifically stabilizes the B56α-PP2A holoenzyme in a fully assembled, active state to dephosphorylate selective substrates, such as its well-known oncogenic target, c-Myc. Our 3.6 Å structure identifies molecular interactions between DT-061 and all three PP2A subunits that prevent dissociation of the active enzyme and highlight inherent mechanisms of PP2A complex assembly. Thus, our findings provide fundamental insights into PP2A complex assembly and regulation, identify a unique interfacial stabilizing mode of action for therapeutic targeting, and aid in the development of phosphatase-based therapeutics tailored against disease specific phospho-protein targets.
Keywords: B56α; PP2A; SMAP; c-Myc; cancer; cryo-EM; phosphatase; small molecule.
Copyright © 2020 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of Interests The Icahn School of Medicine at Mount Sinai has filed patents covering composition of matter on the small molecules disclosed herein for the treatment of human cancer and other diseases (International Application numbers PCT/US15/19770 and PCT/US15/19764 and US Patent number US 9,540,358 B2). Mount Sinai is actively seeking commercial partners for the further development of the technology. G.N. has a financial interest in the commercialization of the technology. RAPPTA Therapeutics has licensed the cryo-EM coordinates for the clinical and commercial development of novel series of small molecule PP2A activators from the University of Michigan (G.N.) and Case Western Reserve University (D.T.). G.N. and D.T. have an ownership interest in RAPPTA Therapeutics. D.L.B. functions as a SAB member for RAPPTA. The Medical University of Vienna, on behalf of E.O., is filing a patent on the PP2A methyl-C subunit specific monoclonal antibody 7C10 disclosed herein for the diagnostic use of 7C10.
Figures
Comment in
-
Piecing Together a Broken Tumor Suppressor Phosphatase for Cancer Therapy.Cell. 2020 Apr 30;181(3):514-517. doi: 10.1016/j.cell.2020.04.005. Cell. 2020. PMID: 32359434
-
Selective stabilization supports phosphatase targeting in cancer.Nat Rev Drug Discov. 2020 Jun;19(6):387. doi: 10.1038/d41573-020-00091-3. Nat Rev Drug Discov. 2020. PMID: 32393817 No abstract available.
-
Select Stabilization of a Tumor-Suppressive PP2A Heterotrimer.Trends Pharmacol Sci. 2020 Sep;41(9):595-597. doi: 10.1016/j.tips.2020.06.008. Epub 2020 Jul 2. Trends Pharmacol Sci. 2020. PMID: 32624198 Free PMC article.
References
-
- Adams PD, Afonine PV, Bunkóczi G, Chen VB, Davis IW, Echols N, Headd JJ, Hung LW, Kapral GJ, Grosse-Kunstleve RW, McCoy AJ, Moriarty NW, Oeffner R, Read RJ, Richardson DC, Richardson JS, Terwilliger TC, Zwart PH, 2010. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr 10.1107/S0907444909052925 - DOI - PMC - PubMed
-
- Allen-Petersen BL, Risom T, Feng Z, Wang Z, Jenny ZP, Thoma MC, Pelz KR, Morton JP, Sansom OJ, Lopez CD, Sheppard B, Christensen DJ, Ohlmeyer M, Narla G, Sears RC, 2018. Activation of PP2A and inhibition of mTOR synergistically reduce MYC signaling and decrease tumor growth in pancreatic ductal adenocarcinoma. Cancer Res 10.1158/0008-5472.CAN-18-0717 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
