

Physicochemical Characterisation of KEIF-The Intrinsically Disordered N-Terminal Region of Magnesium Transporter A
- PMID: 32316569
- PMCID: PMC7226168
- DOI: 10.3390/biom10040623
Physicochemical Characterisation of KEIF-The Intrinsically Disordered N-Terminal Region of Magnesium Transporter A
Abstract
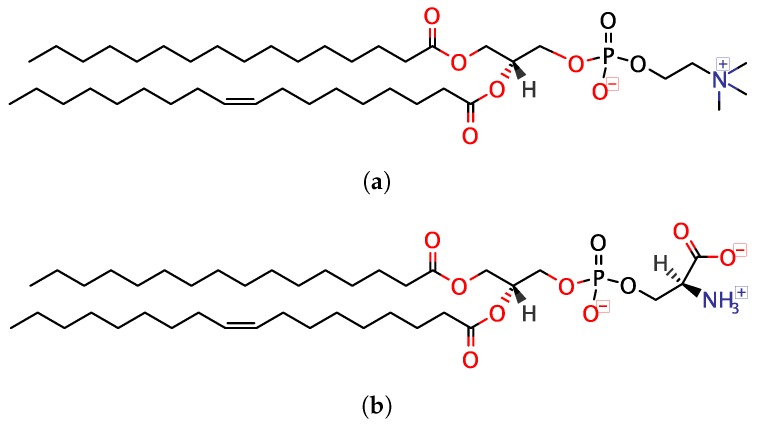
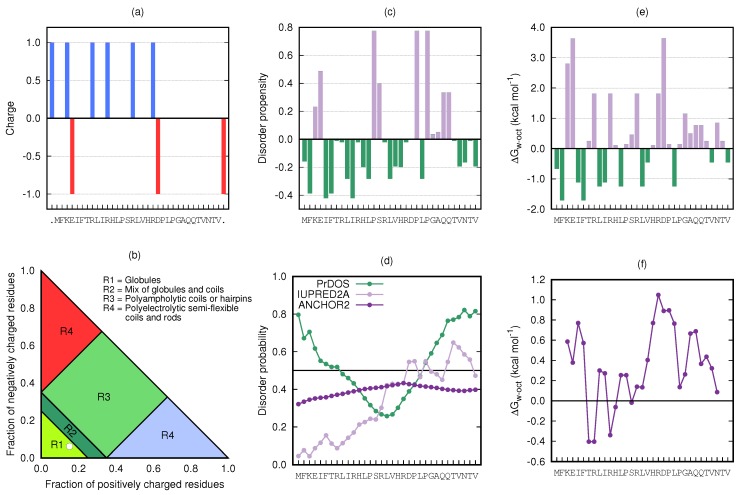
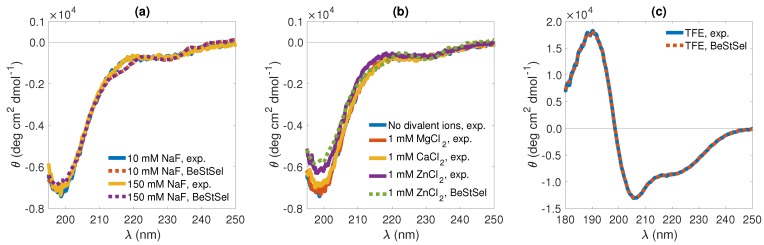
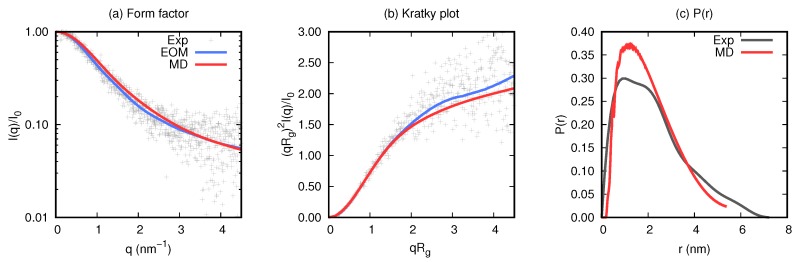
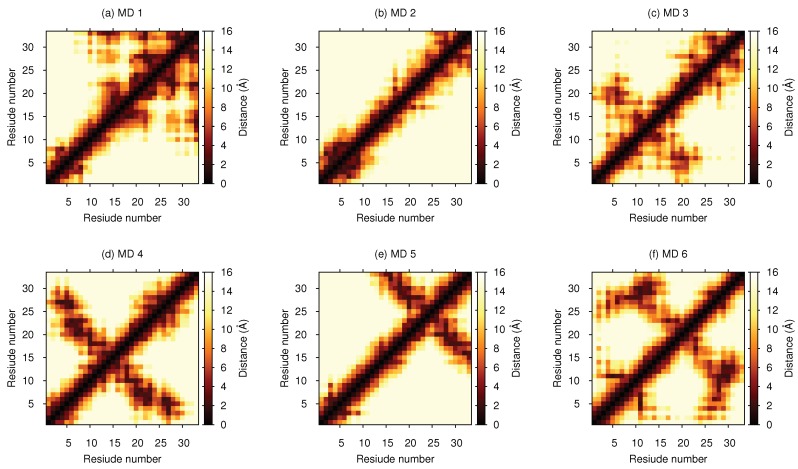
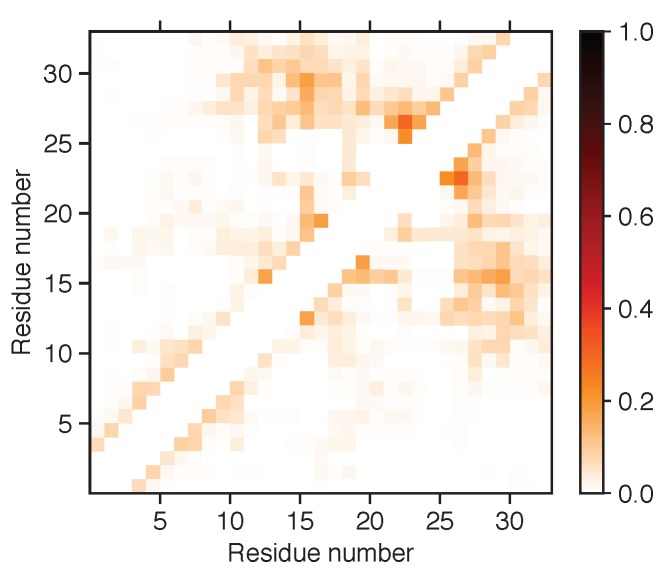
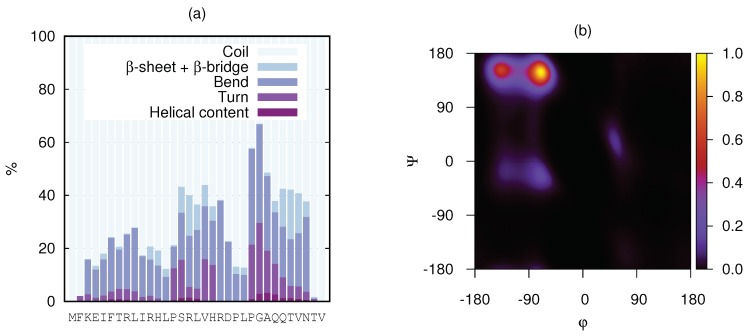
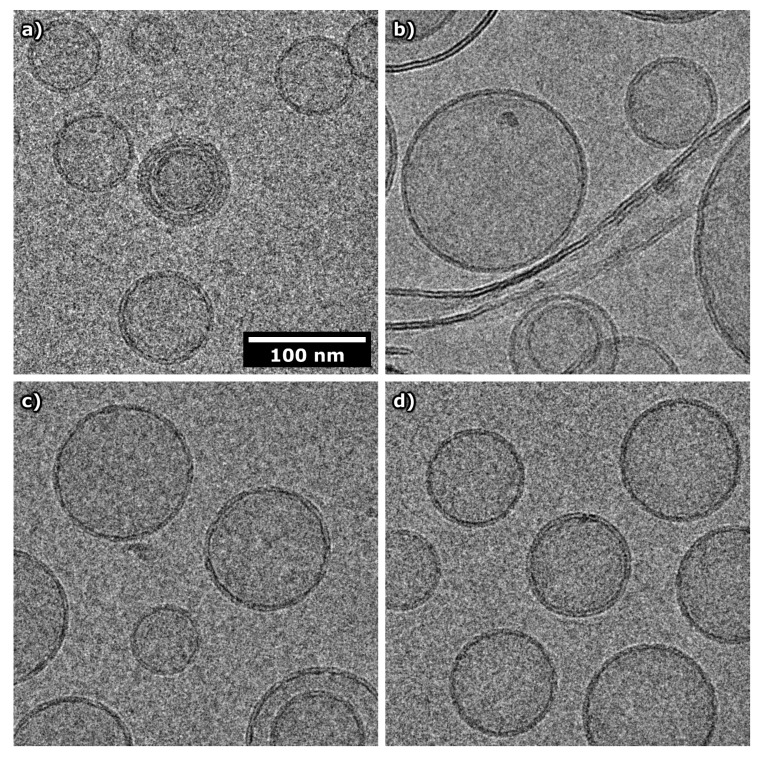
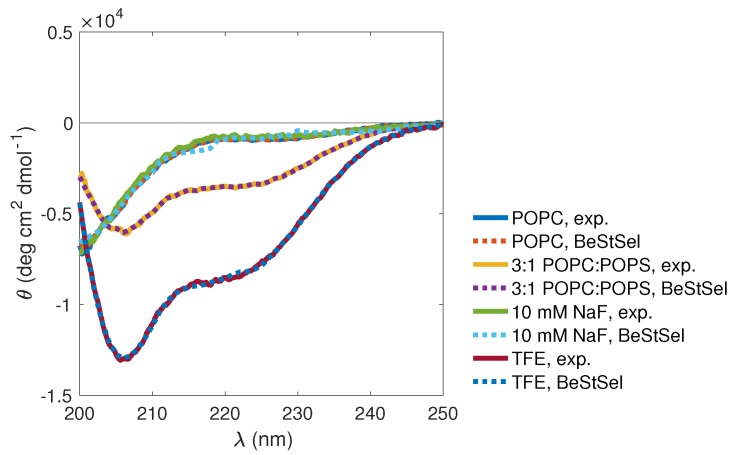
Magnesium transporter A (MgtA) is an active transporter responsible for importing magnesium ions into the cytoplasm of prokaryotic cells. This study focuses on the peptide corresponding to the intrinsically disordered N-terminal region of MgtA, referred to as KEIF. Primary-structure and bioinformatic analyses were performed, followed by studies of the undisturbed single chain using a combination of techniques including small-angle X-ray scattering, circular dichroism spectroscopy, and atomistic molecular-dynamics simulations. Moreover, interactions with large unilamellar vesicles were investigated by using dynamic light scattering, laser Doppler velocimetry, cryogenic transmission electron microscopy, and circular dichroism spectroscopy. KEIF was confirmed to be intrinsically disordered in aqueous solution, although extended and containing little β -structure and possibly PPII structure. An increase of helical content was observed in organic solvent, and a similar effect was also seen in aqueous solution containing anionic vesicles. Interactions of cationic KEIF with anionic vesicles led to the hypothesis that KEIF adsorbs to the vesicle surface through electrostatic and entropic driving forces. Considering this, there is a possibility that the biological role of KEIF is to anchor MgtA in the cell membrane, although further investigation is needed to confirm this hypothesis.
Keywords: circular dichroism spectroscopy; cryogenic transmission electron microscopy; intrinsically disordered proteins; magnesium transporter; membrane proteins; molecular-dynamics simulations; protein–vesicle interactions; secondary structure.; small-angle X-ray scattering.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
