

Host Cathelicidin Exacerbates Group B Streptococcus Urinary Tract Infection
- PMID: 32321824
- PMCID: PMC7178553
- DOI: 10.1128/mSphere.00932-19
Host Cathelicidin Exacerbates Group B Streptococcus Urinary Tract Infection
Abstract
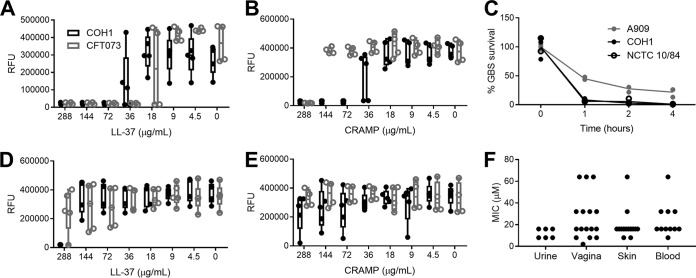
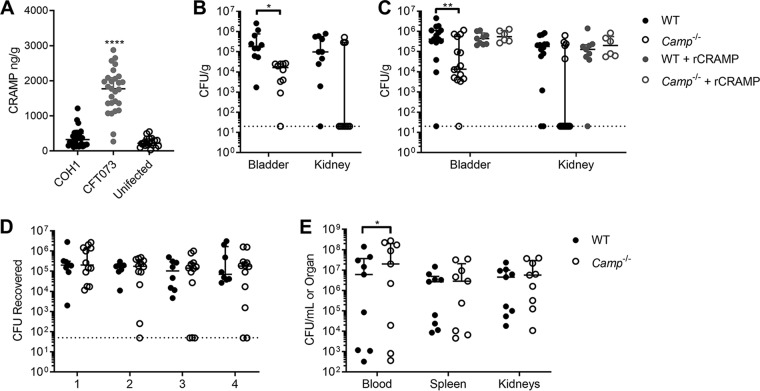
Group B Streptococcus (GBS) causes frequent urinary tract infection (UTI) in susceptible populations, including individuals with type 2 diabetes and pregnant women; however, specific host factors responsible for increased GBS susceptibility in these populations are not well characterized. Here, we investigate cathelicidin, a cationic antimicrobial peptide, known to be critical for defense during UTI with uropathogenic Escherichia coli (UPEC). We observed a loss of antimicrobial activity of human and mouse cathelicidins against GBS and UPEC in synthetic urine and no evidence for increased cathelicidin resistance in GBS urinary isolates. Furthermore, we found that GBS degrades cathelicidin in a protease-dependent manner. Surprisingly, in a UTI model, cathelicidin-deficient (Camp-/-) mice showed decreased GBS burdens and mast cell recruitment in the bladder compared to levels in wild-type (WT) mice. Pharmacologic inhibition of mast cells reduced GBS burdens and histamine release in WT but not Camp-/- mice. Streptozotocin-induced diabetic mice had increased bladder cathelicidin production and mast cell recruitment at 24 h postinfection with GBS compared to levels in nondiabetic controls. We propose that cathelicidin is an important immune regulator but ineffective antimicrobial peptide against GBS in urine. Combined, our findings may in part explain the increased frequency of GBS UTI in diabetic and pregnant individuals.IMPORTANCE Certain populations such as diabetic individuals are at increased risk for developing urinary tract infections (UTI), although the underlying reasons for this susceptibility are not fully known. Additionally, diabetics are more likely to become infected with certain types of bacteria, such as group B Streptococcus (GBS). In this study, we find that an antimicrobial peptide called cathelicidin, which is thought to protect the bladder from infection, is ineffective in controlling GBS and alters the type of immune cells that migrate to the bladder during infection. Using a mouse model of diabetes, we observe that diabetic mice are more susceptible to GBS infection even though they also have more infiltrating immune cells and increased production of cathelicidin. Taken together, our findings identify this antimicrobial peptide as a potential contributor to increased susceptibility of diabetic individuals to GBS UTI.
Keywords: cathelicidin; group B Streptococcus; innate immunity; mast cell; urinary tract infection.
Copyright © 2020 Patras et al.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
