

The gut-brain axis mediates sugar preference
- PMID: 32322067
- PMCID: PMC7185044
- DOI: 10.1038/s41586-020-2199-7
The gut-brain axis mediates sugar preference
Abstract
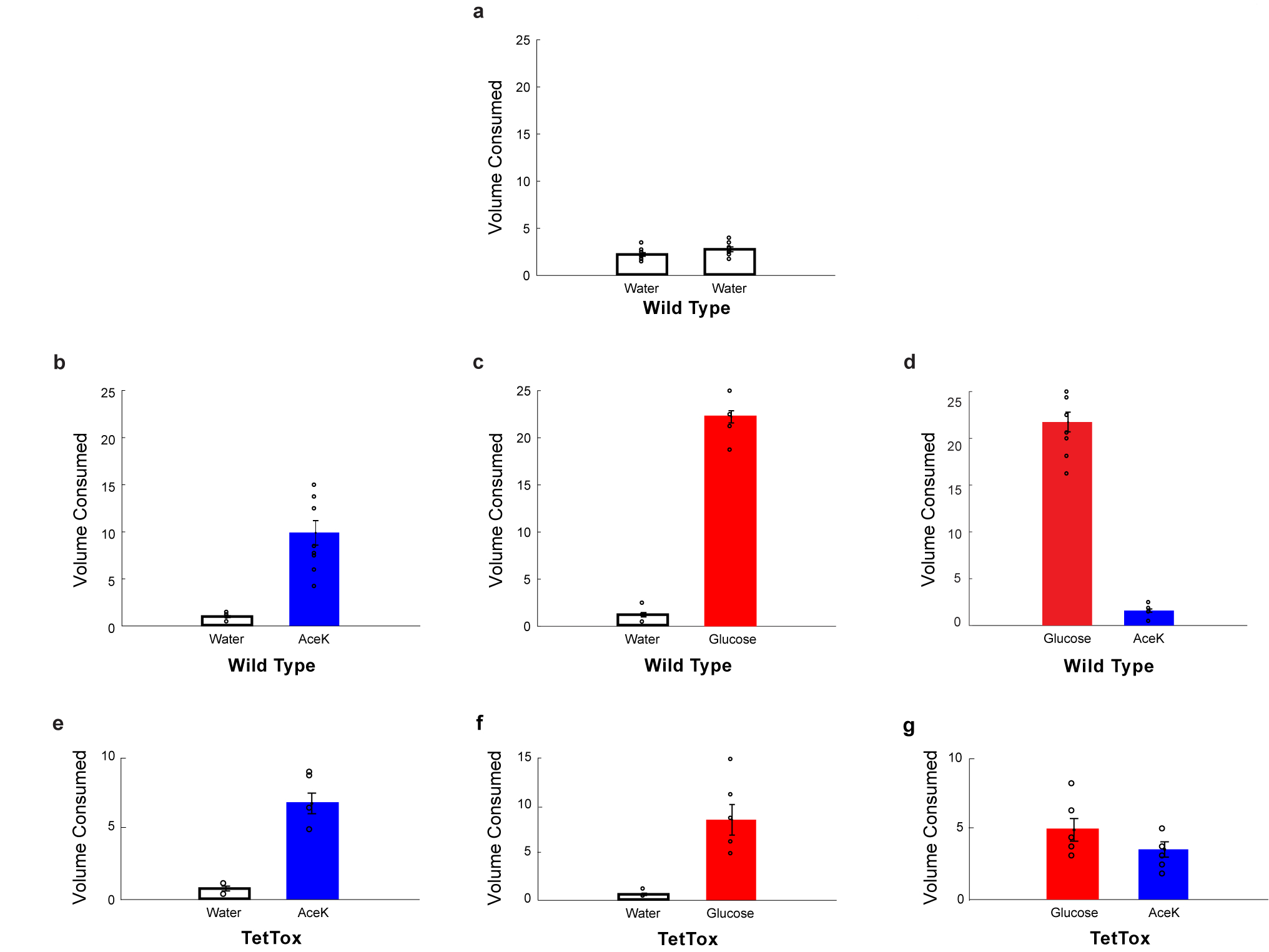
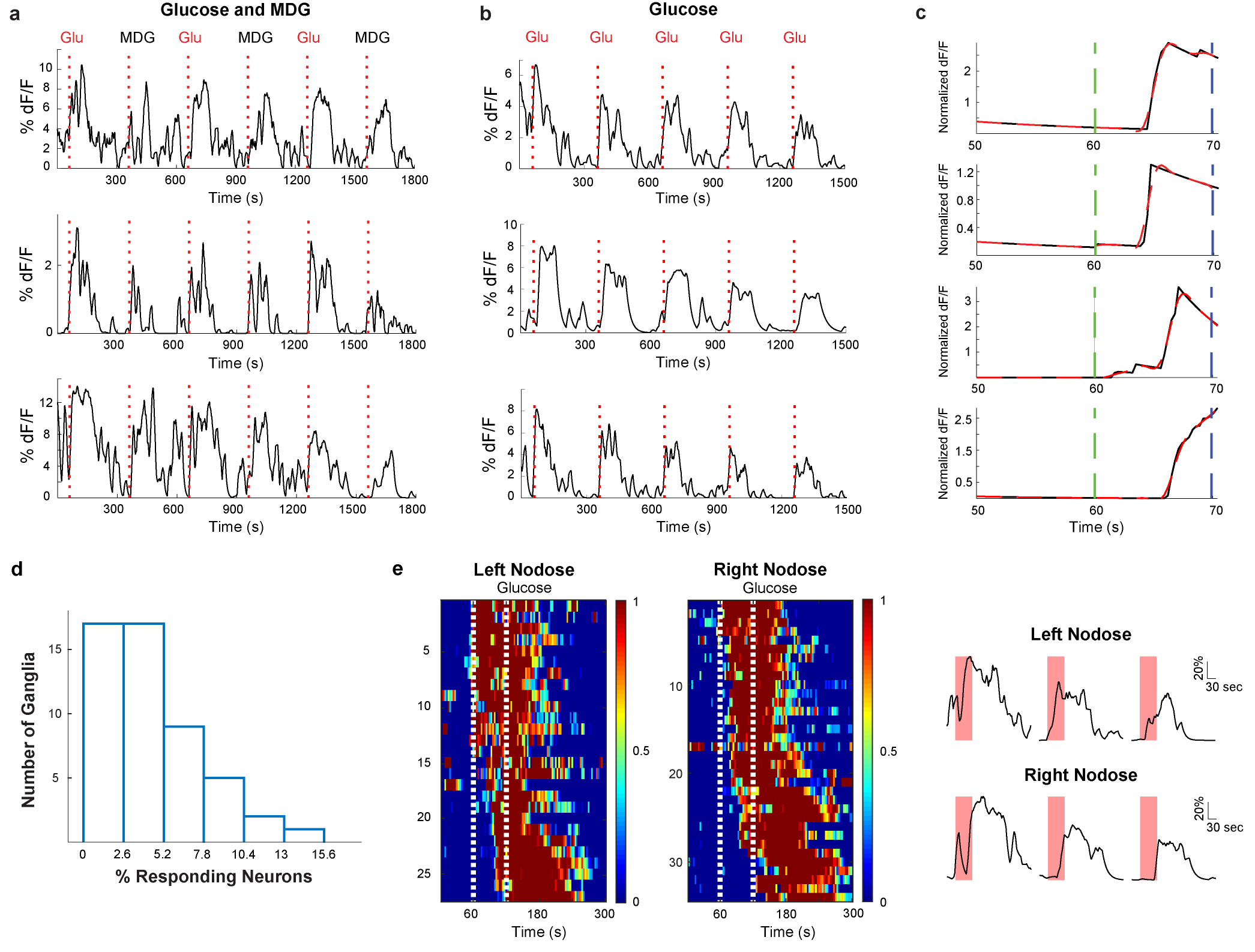
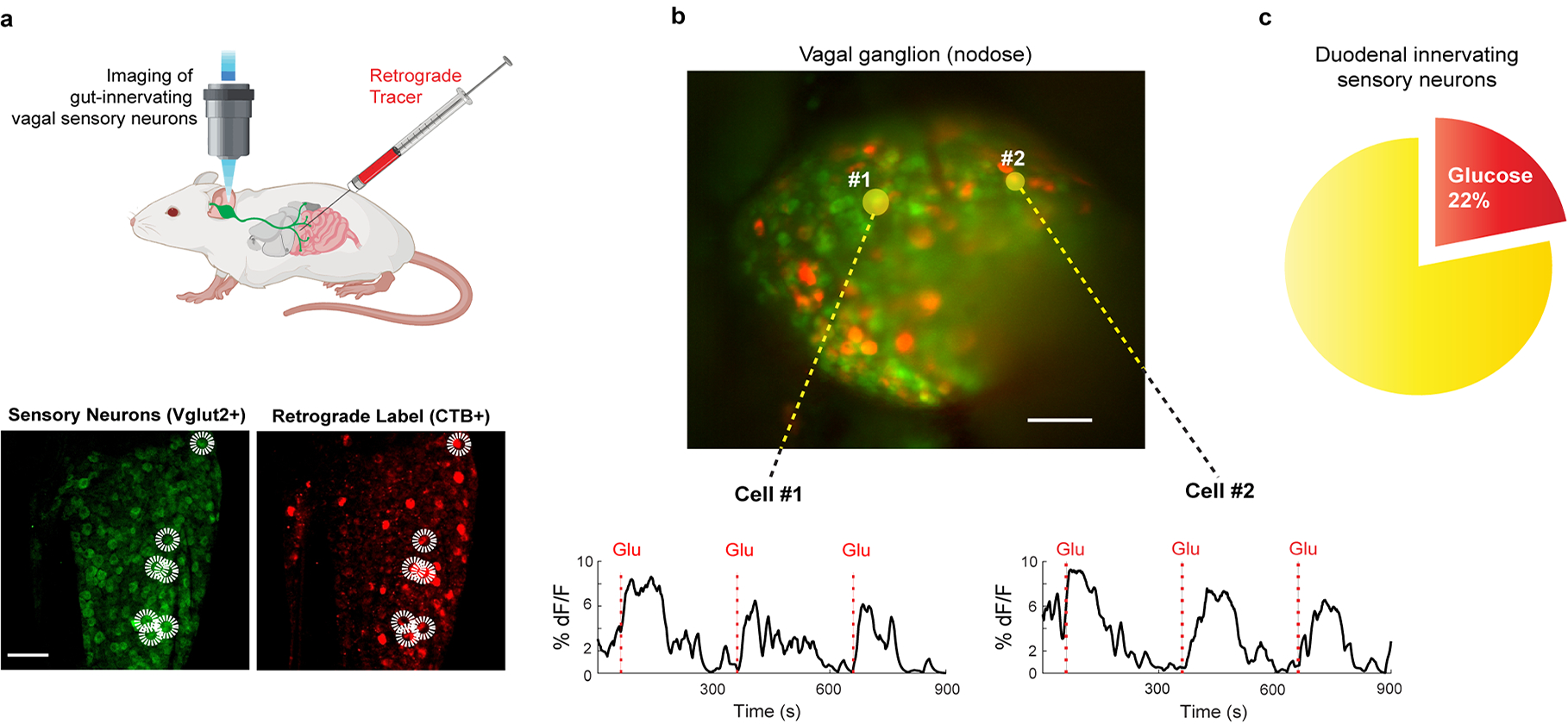
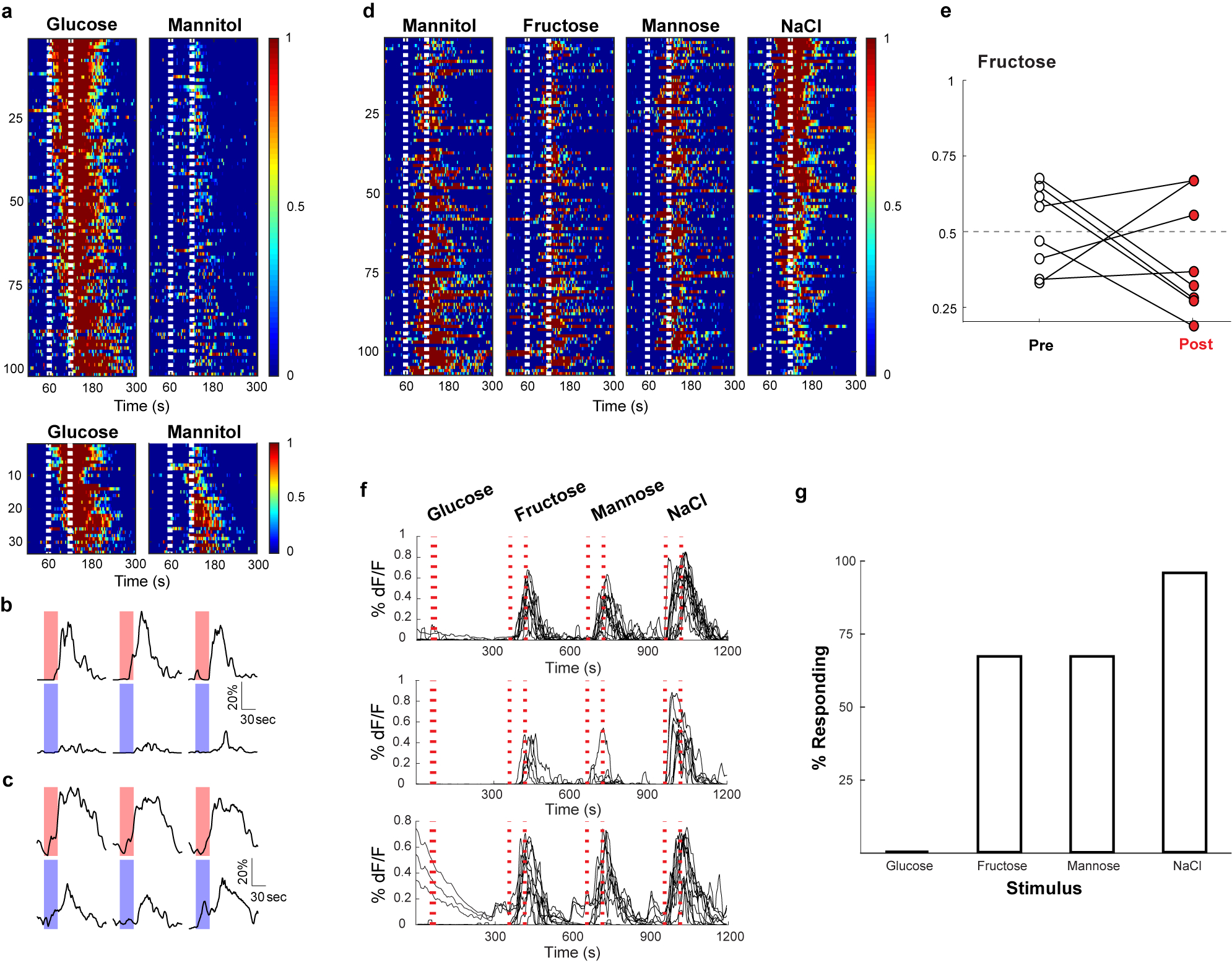
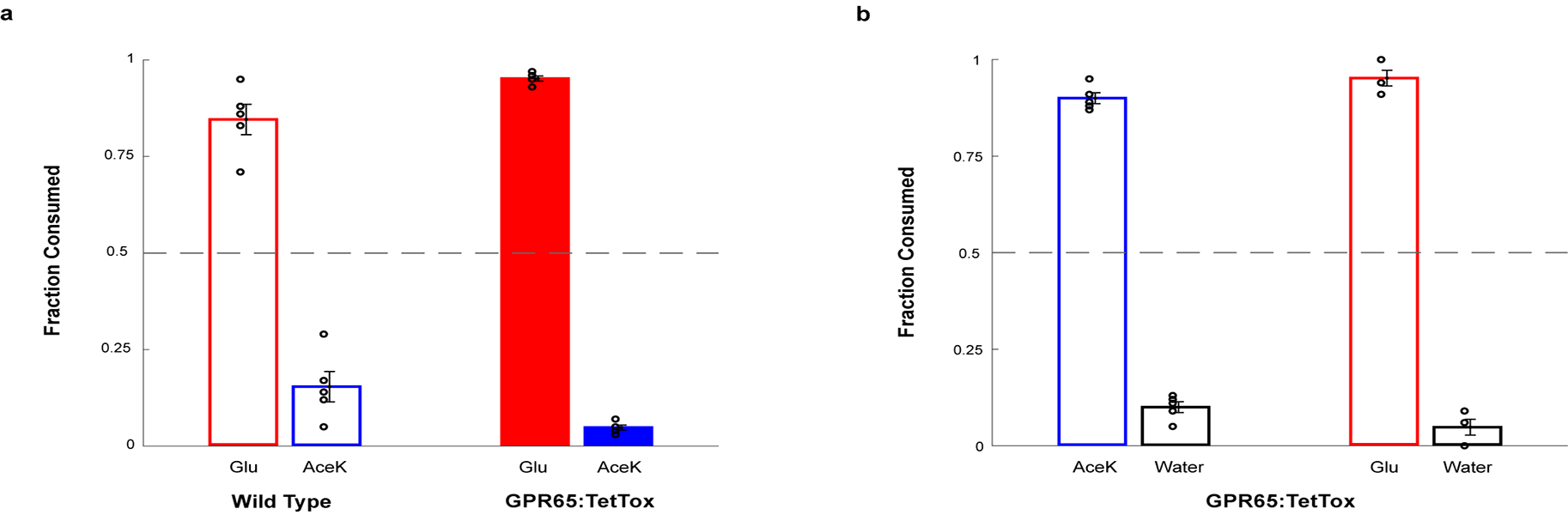
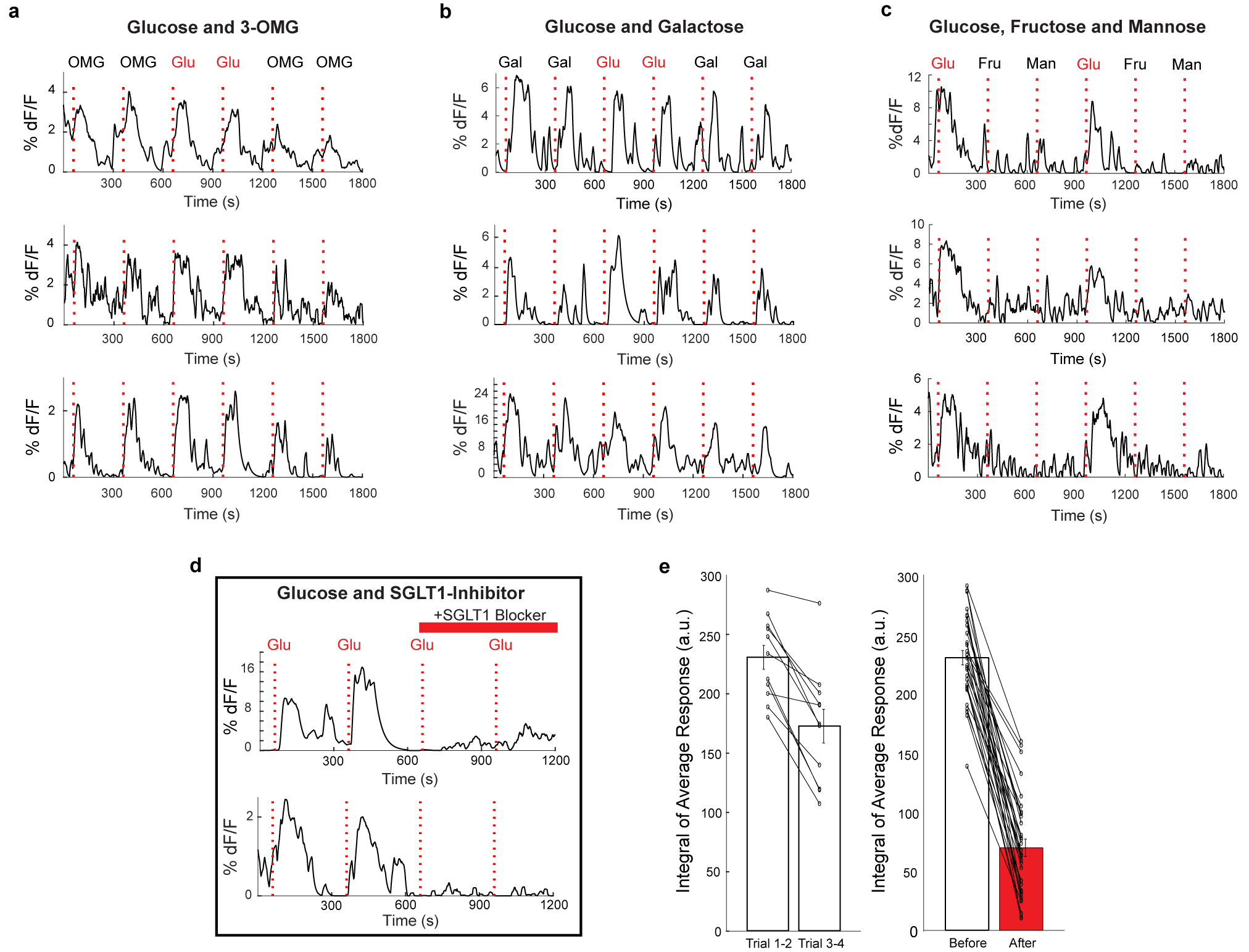
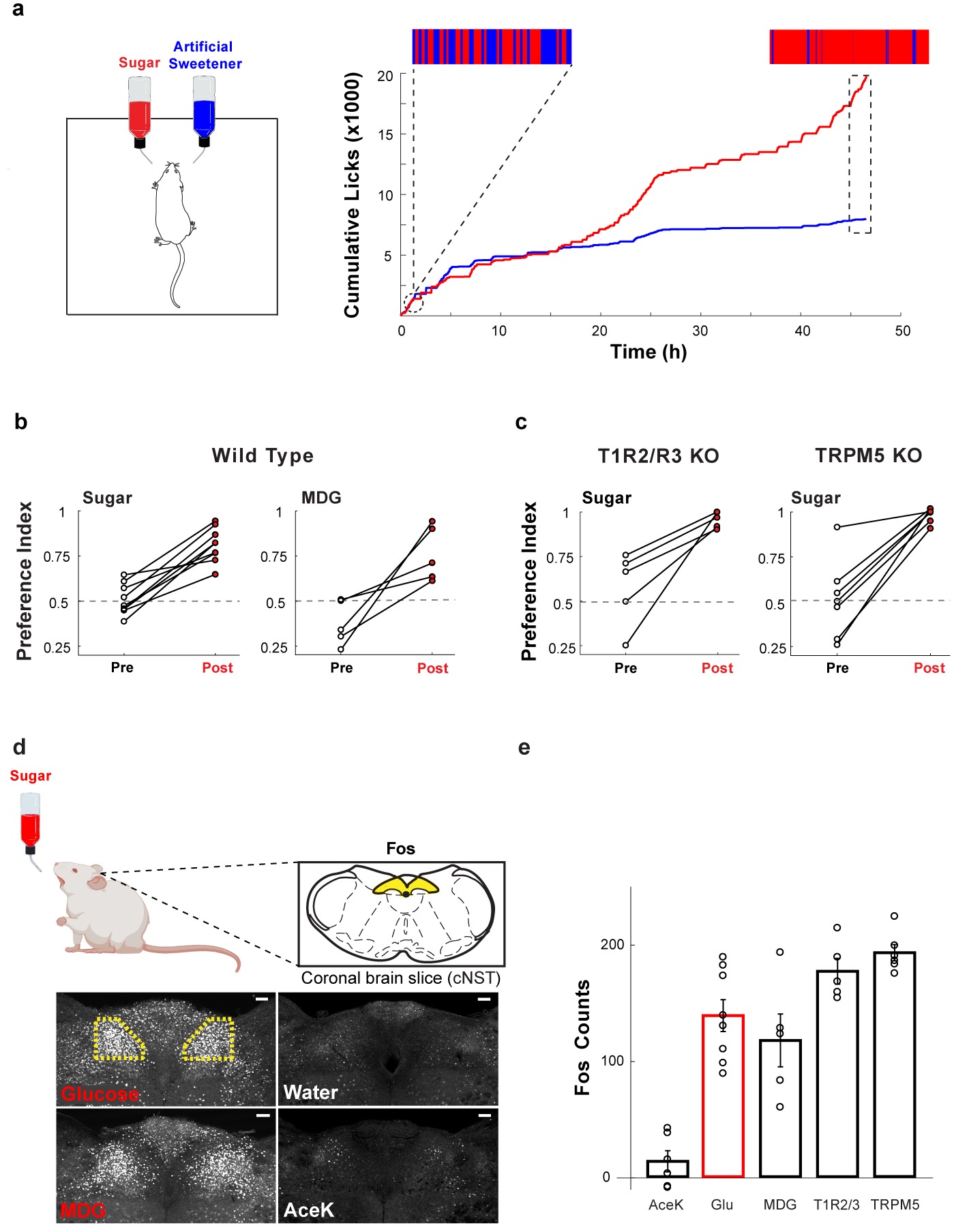
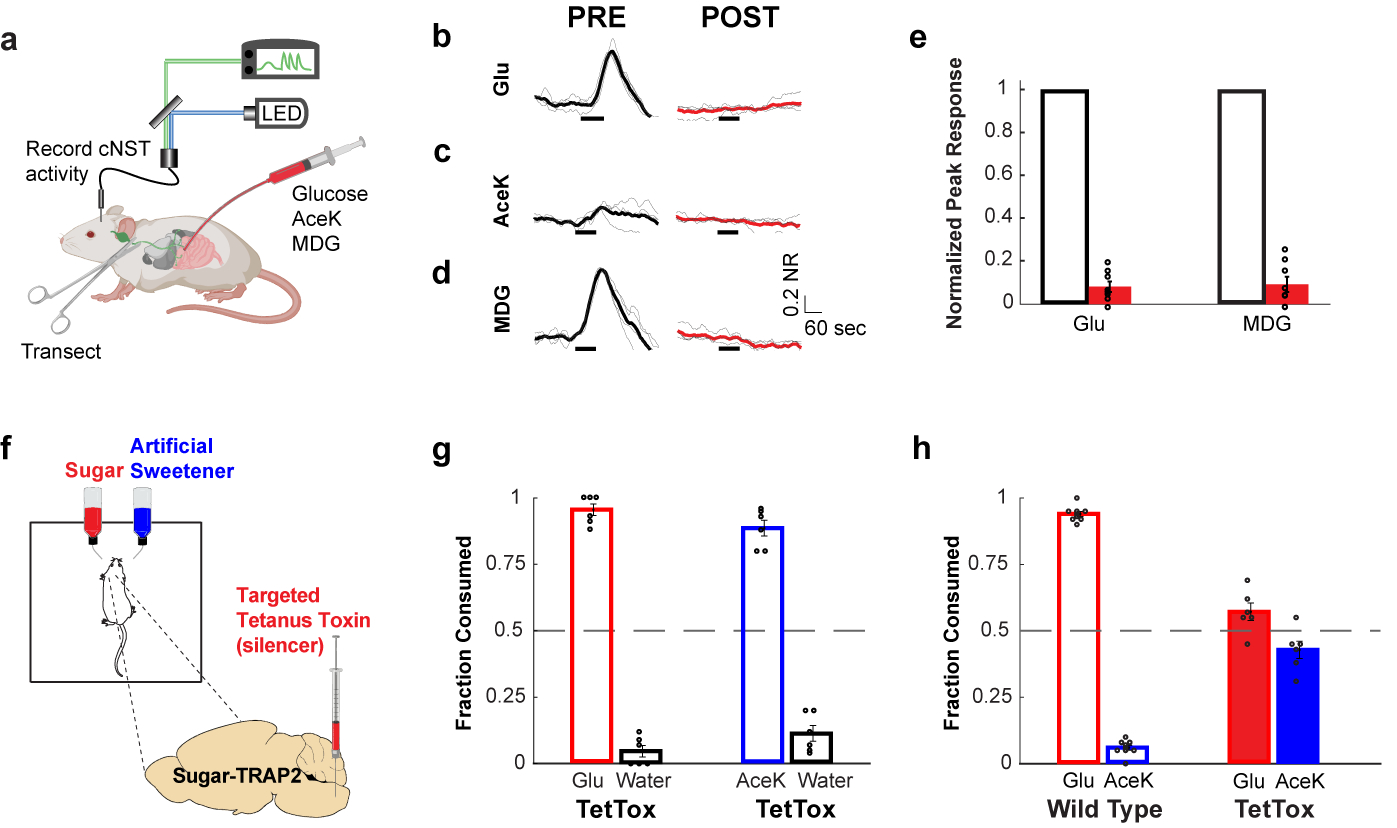
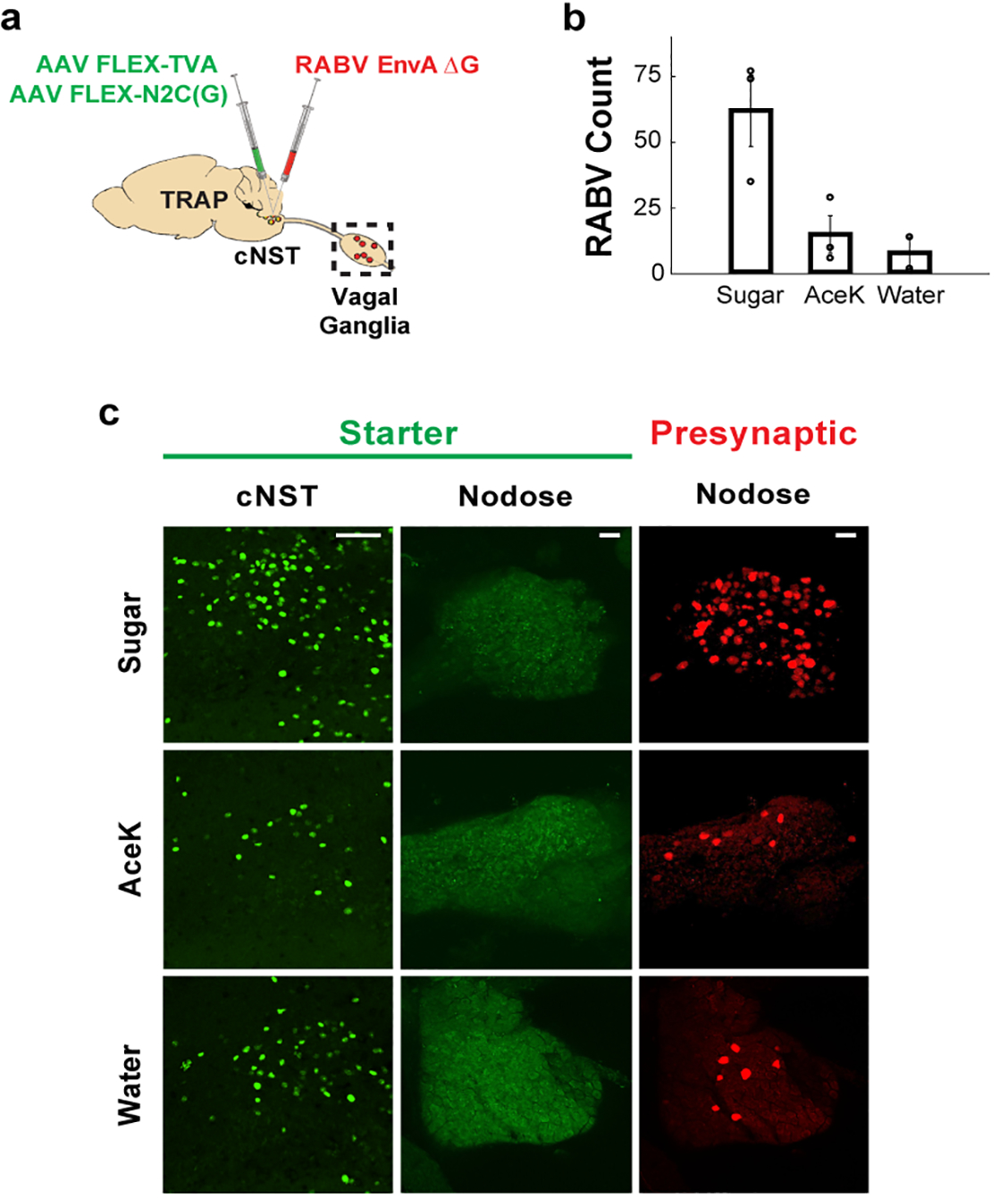
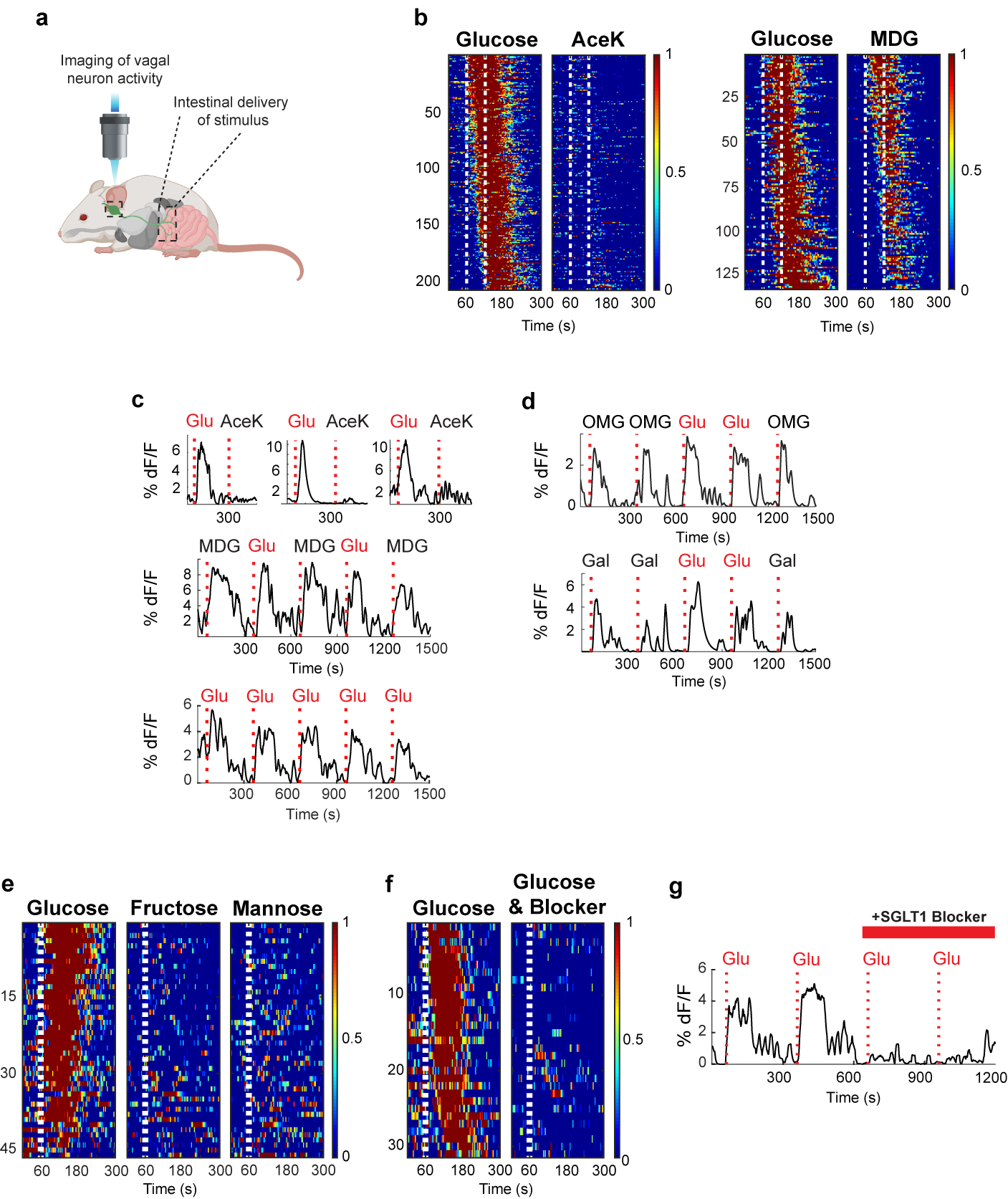
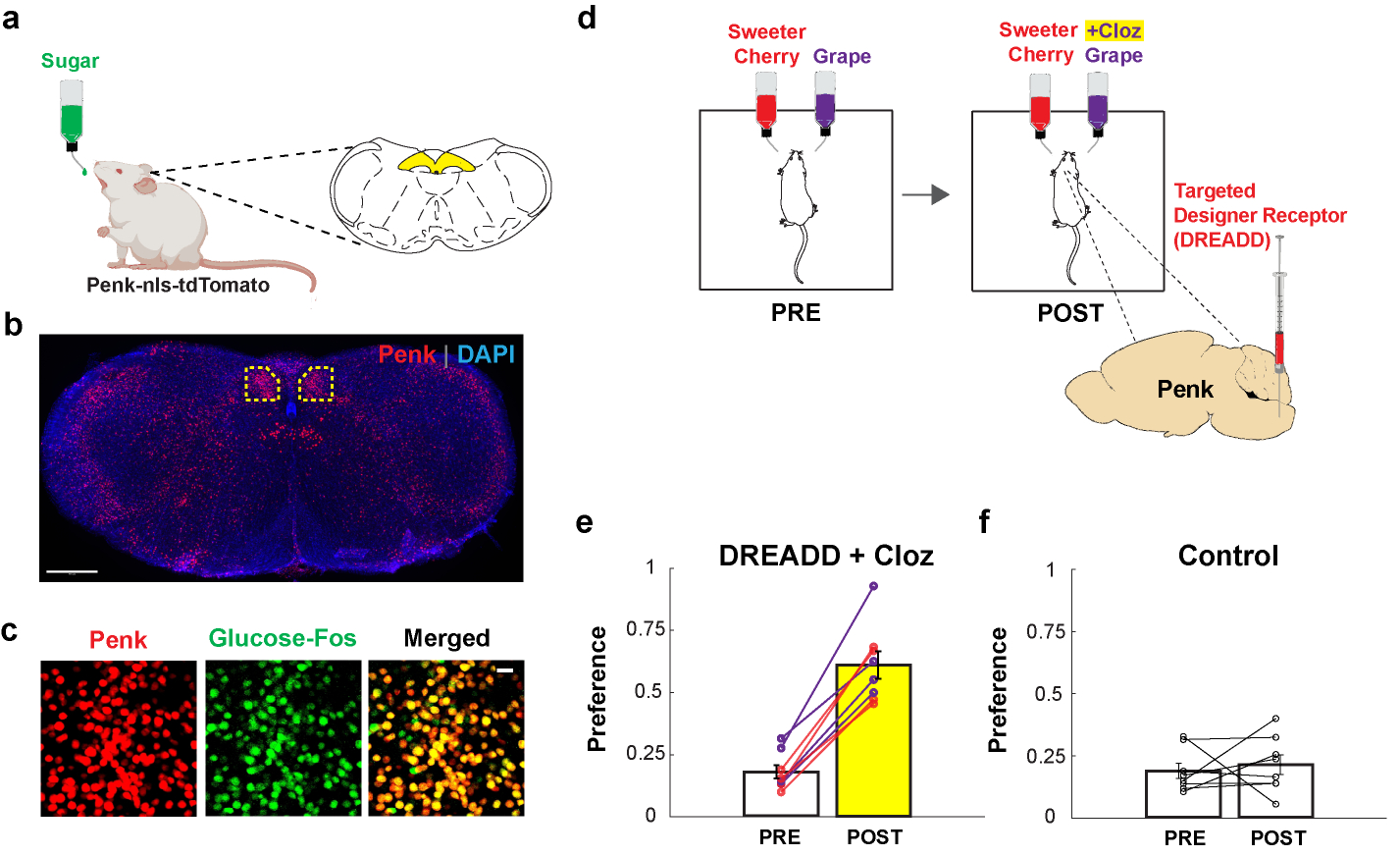
The taste of sugar is one of the most basic sensory percepts for humans and other animals. Animals can develop a strong preference for sugar even if they lack sweet taste receptors, indicating a mechanism independent of taste1-3. Here we examined the neural basis for sugar preference and demonstrate that a population of neurons in the vagal ganglia and brainstem are activated via the gut-brain axis to create preference for sugar. These neurons are stimulated in response to sugar but not artificial sweeteners, and are activated by direct delivery of sugar to the gut. Using functional imaging we monitored activity of the gut-brain axis, and identified the vagal neurons activated by intestinal delivery of glucose. Next, we engineered mice in which synaptic activity in this gut-to-brain circuit was genetically silenced, and prevented the development of behavioural preference for sugar. Moreover, we show that co-opting this circuit by chemogenetic activation can create preferences to otherwise less-preferred stimuli. Together, these findings reveal a gut-to-brain post-ingestive sugar-sensing pathway critical for the development of sugar preference. In addition, they explain the neural basis for differences in the behavioural effects of sweeteners versus sugar, and uncover an essential circuit underlying the highly appetitive effects of sugar.
Conflict of interest statement
Competing Interests
C.S.Z. is a scientific co-founder and advisor of Kallyope.
Figures
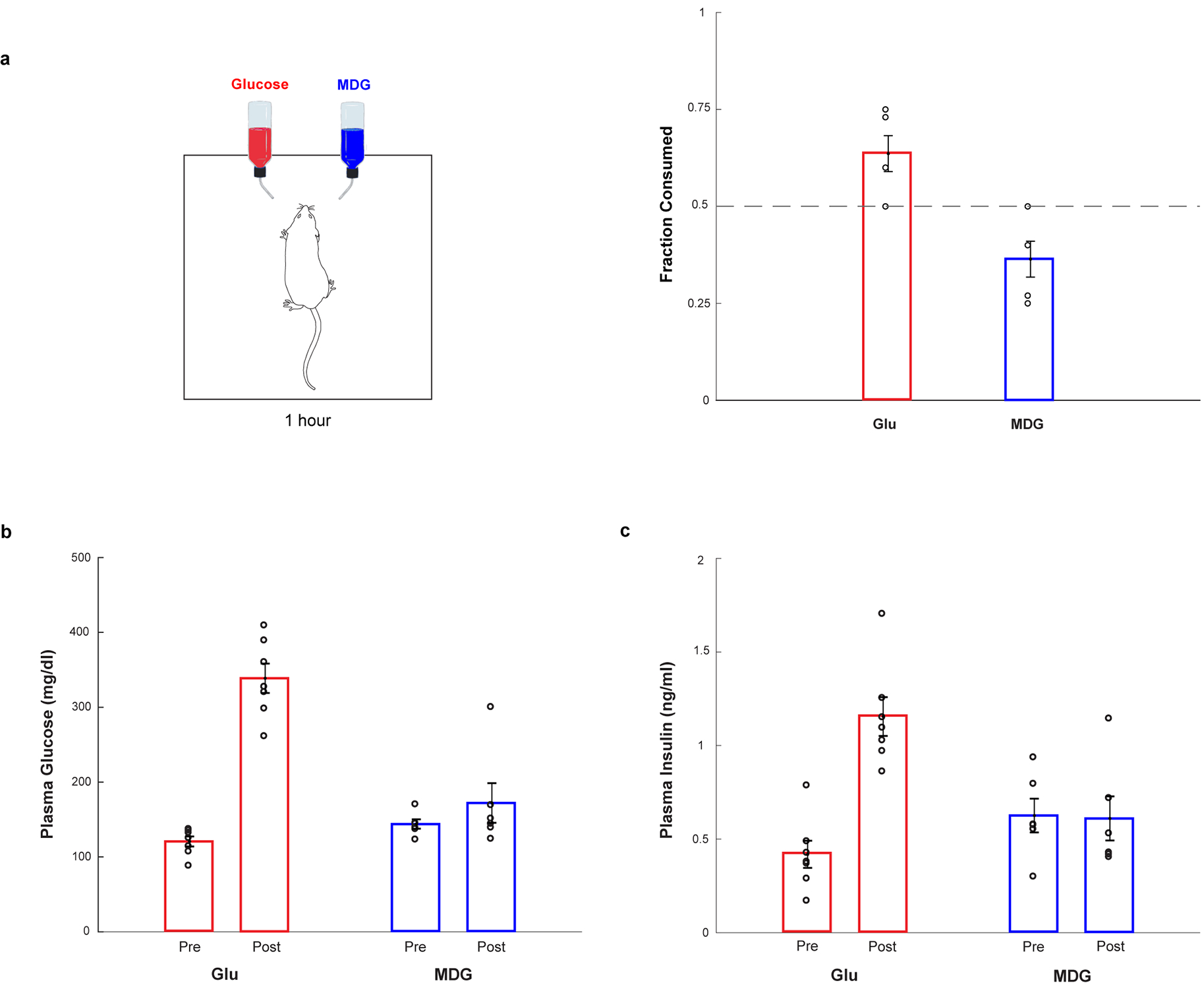
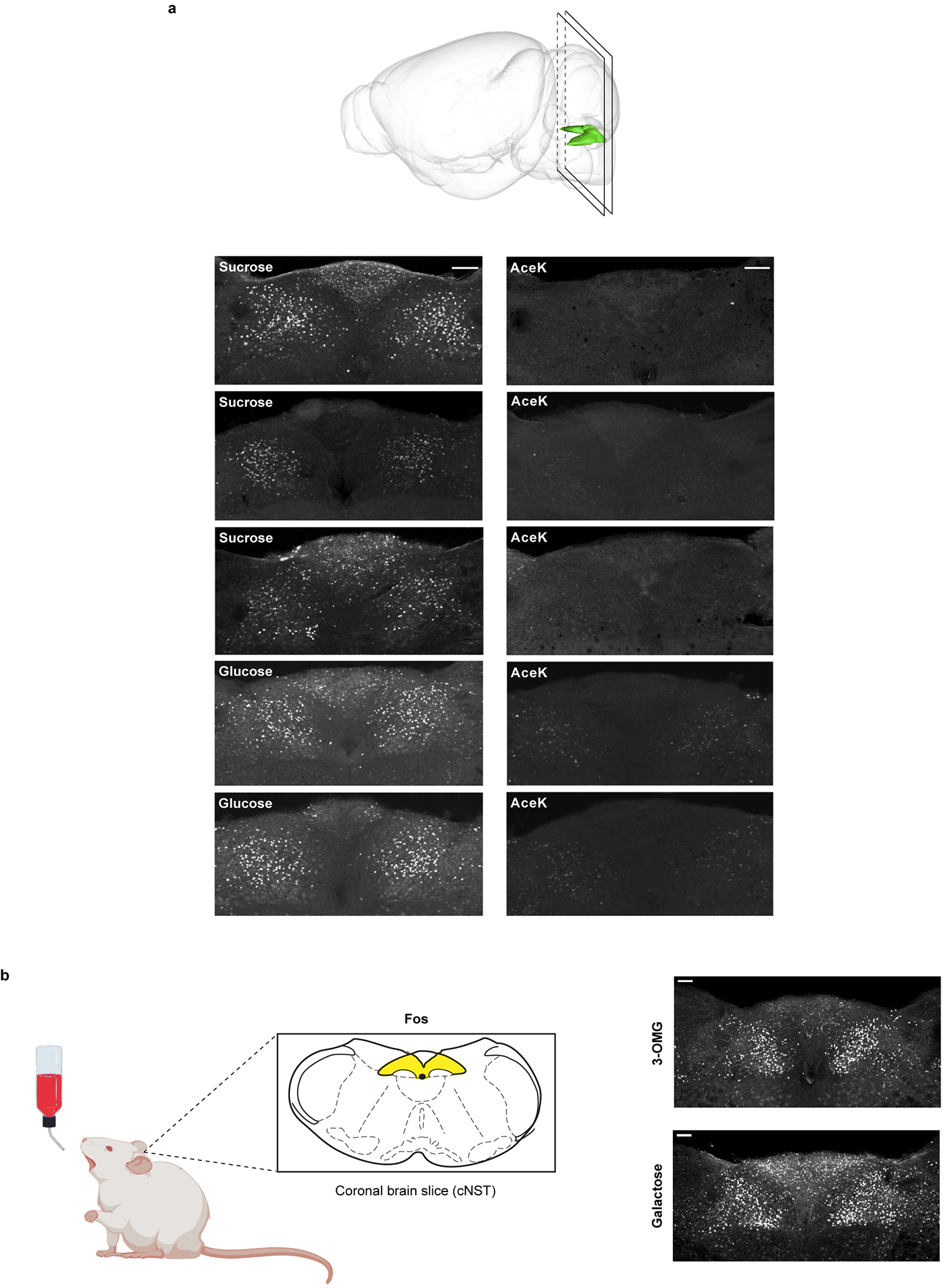
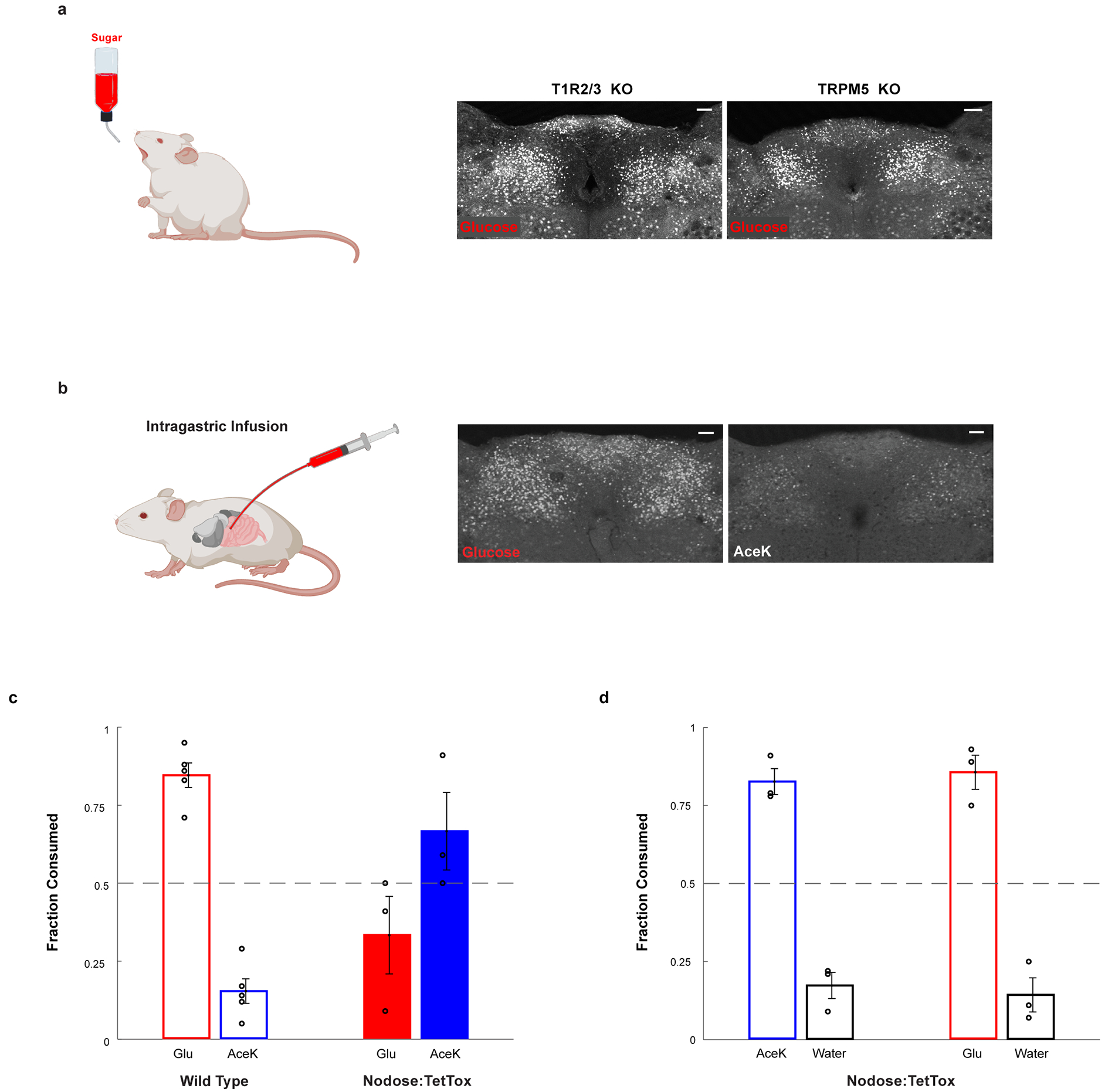
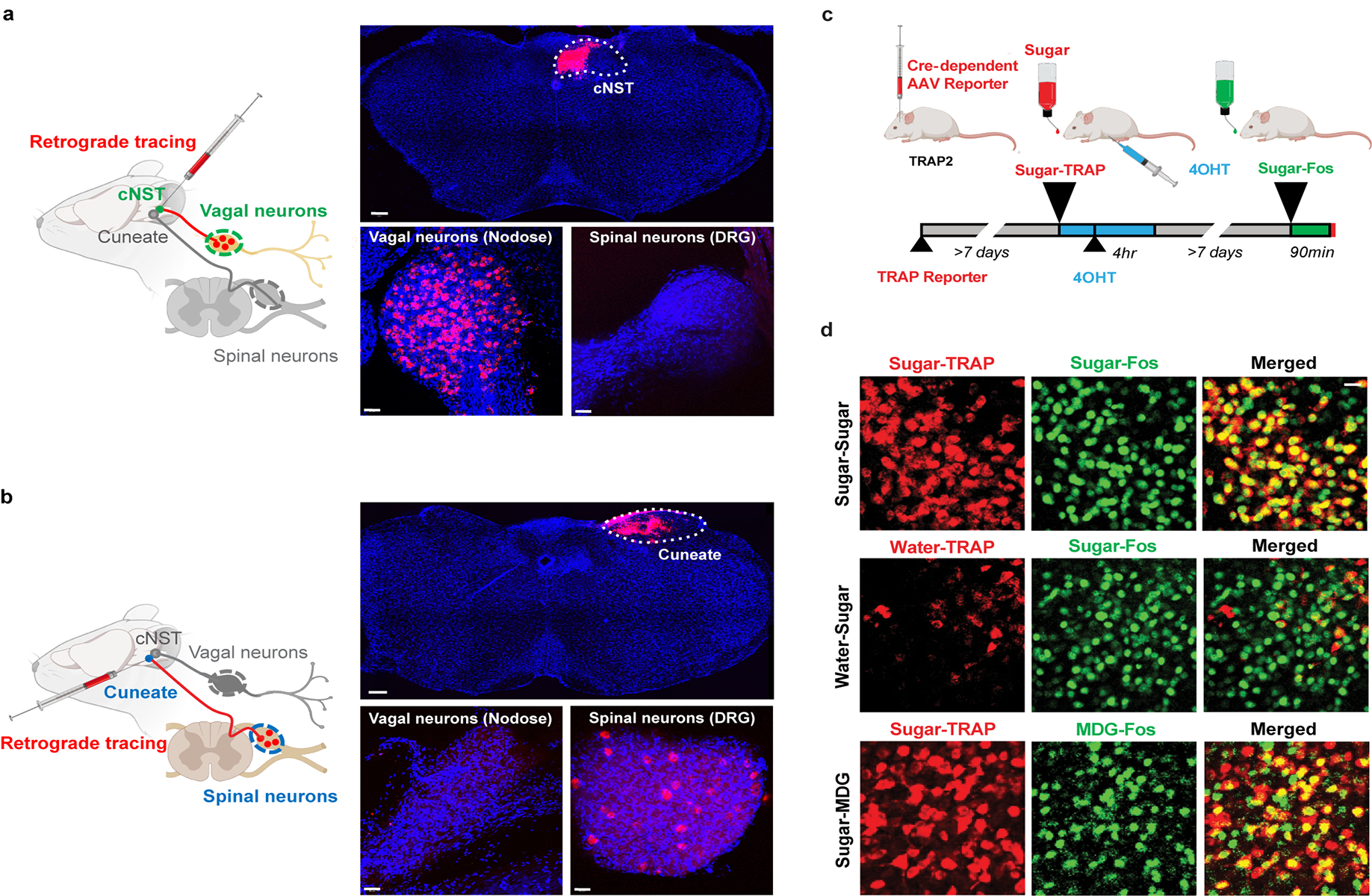
Comment in
-
Sweet signals - gut-brain circuit for sugar identified.Nat Rev Endocrinol. 2020 Jul;16(7):344-345. doi: 10.1038/s41574-020-0366-4. Nat Rev Endocrinol. 2020. PMID: 32346114 No abstract available.
-
A gut instinct for glucose.Nat Rev Neurosci. 2020 Jun;21(6):301. doi: 10.1038/s41583-020-0312-4. Nat Rev Neurosci. 2020. PMID: 32350437 No abstract available.
References
-
- de Araujo IE et al. Food Reward in the Absence of Taste Receptor Signaling. Neuron 57, 930–941 (2008). - PubMed
-
- Elliott Perry. & Elliott P. Production of sugar in the United States and foreign countries. (U.S. Dept. of Agriculture, 1917).
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
