

Oocytes mount a noncanonical DNA damage response involving APC-Cdh1-mediated proteolysis
- PMID: 32328643
- PMCID: PMC7147104
- DOI: 10.1083/jcb.201907213
Oocytes mount a noncanonical DNA damage response involving APC-Cdh1-mediated proteolysis
Abstract
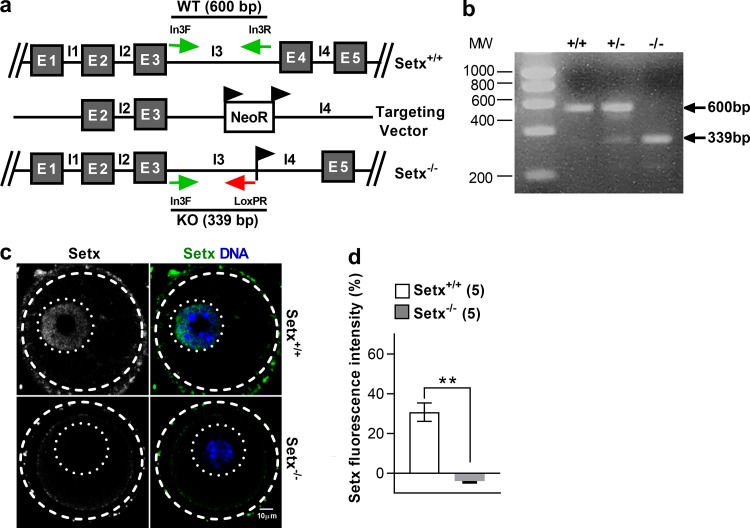
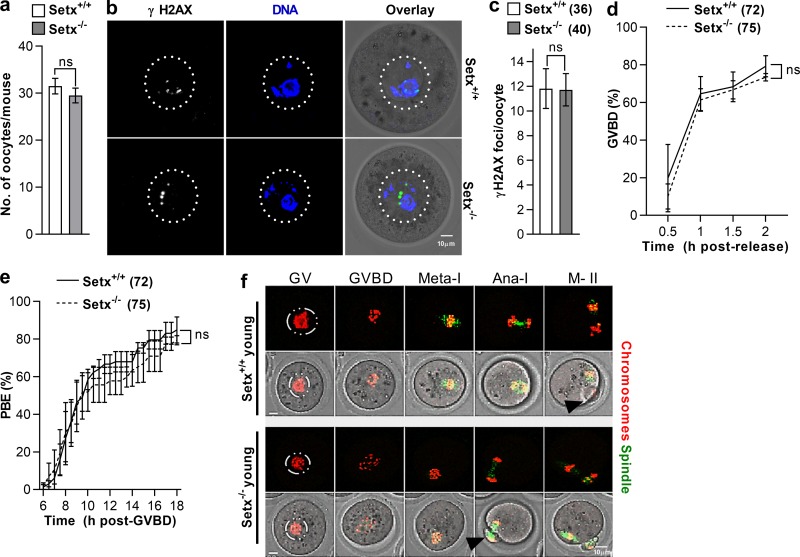
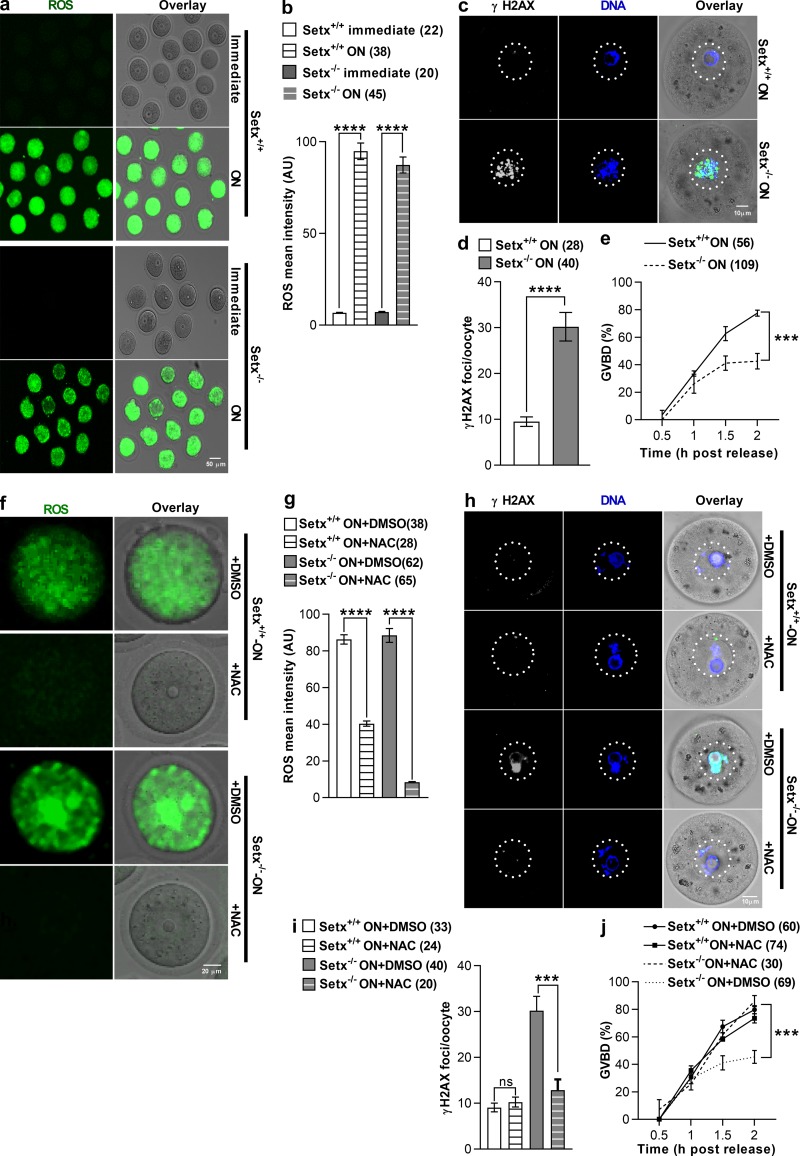
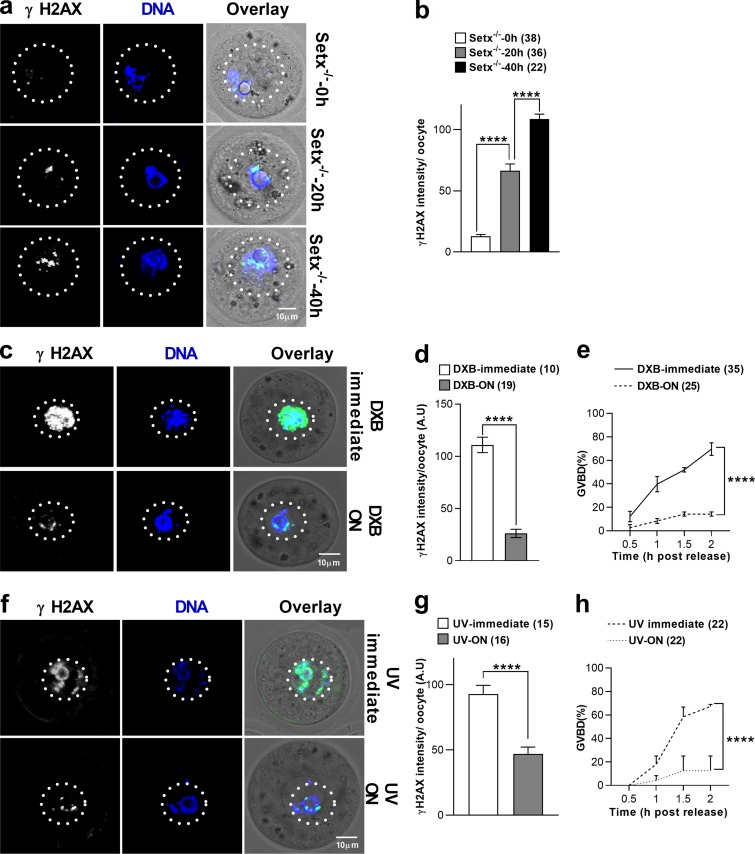
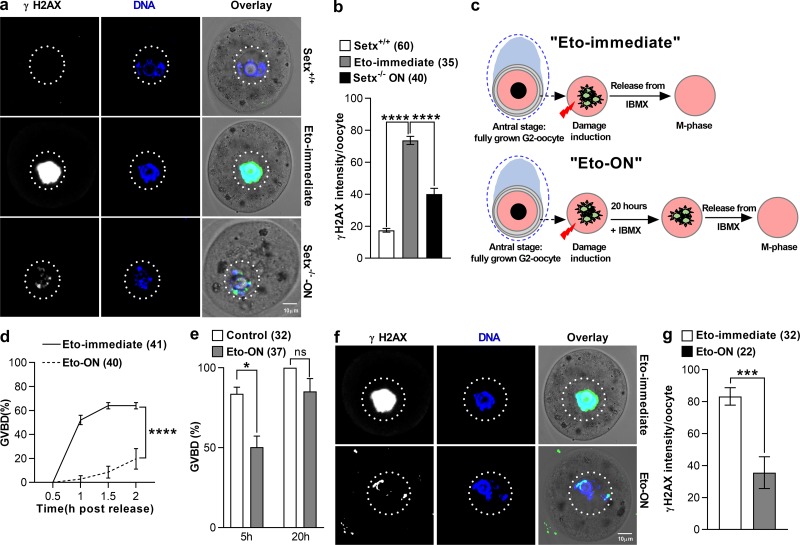
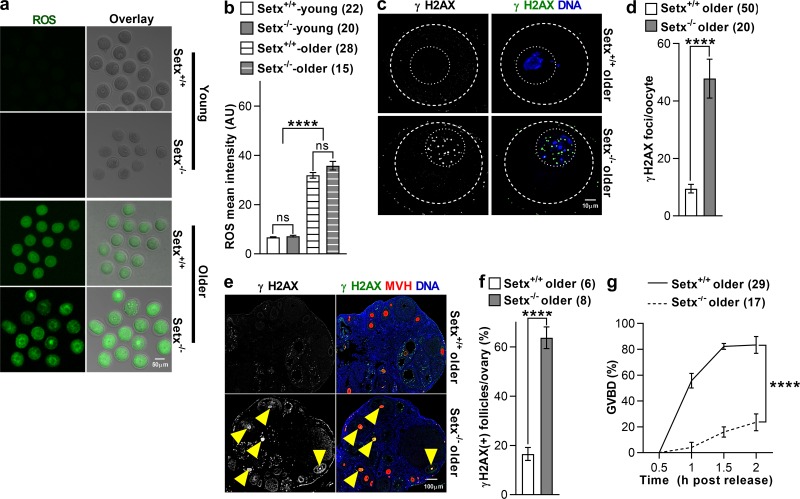
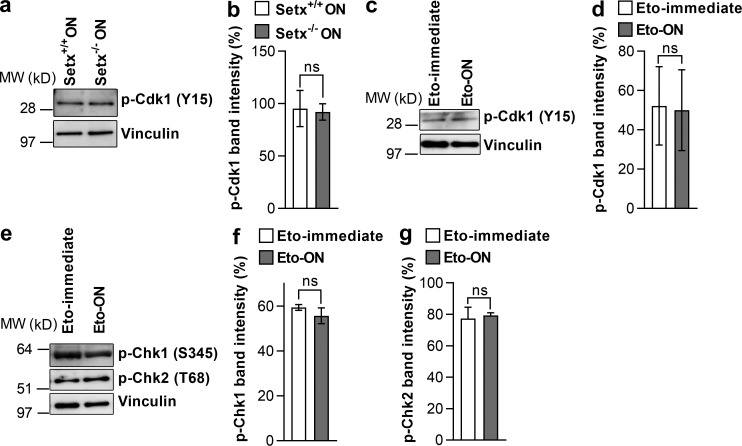
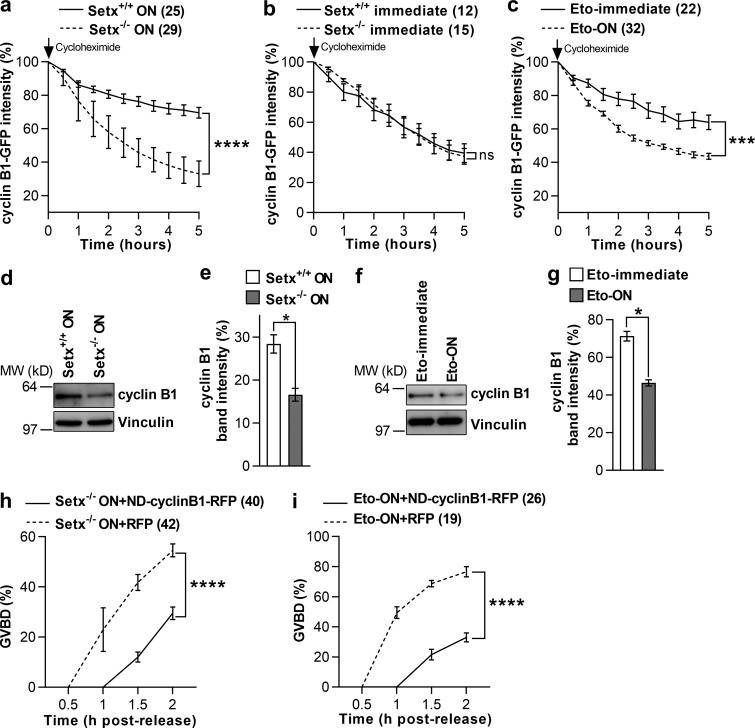
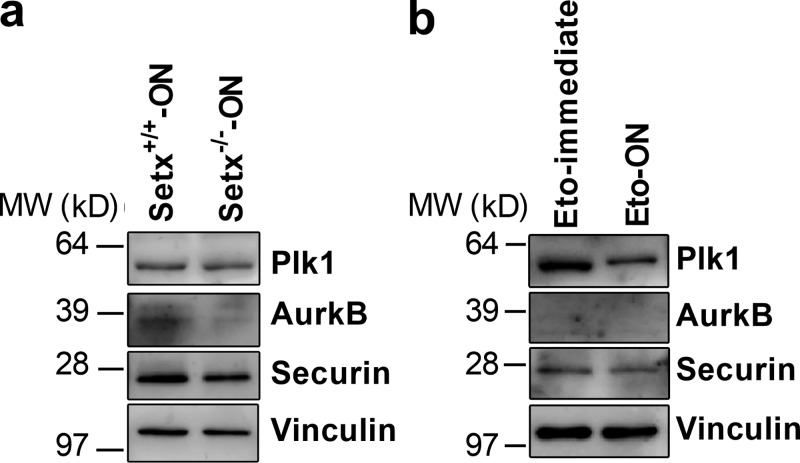
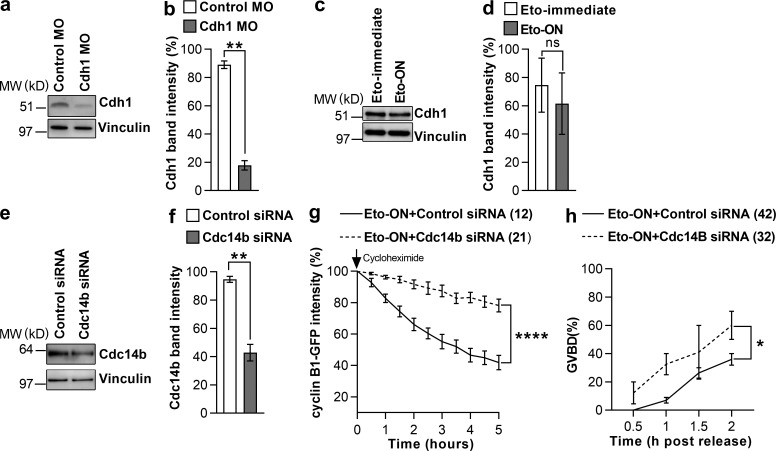
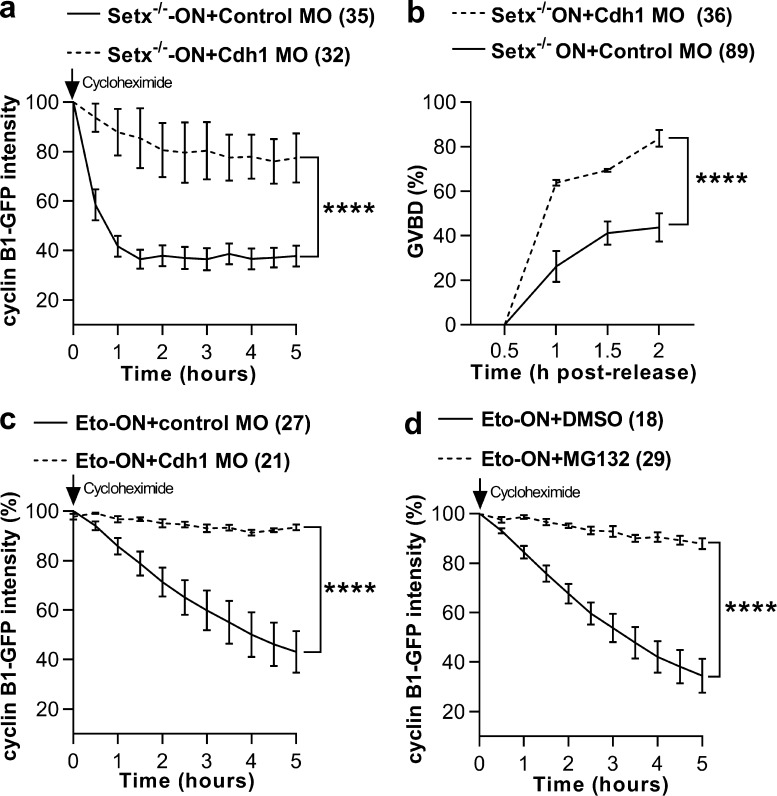
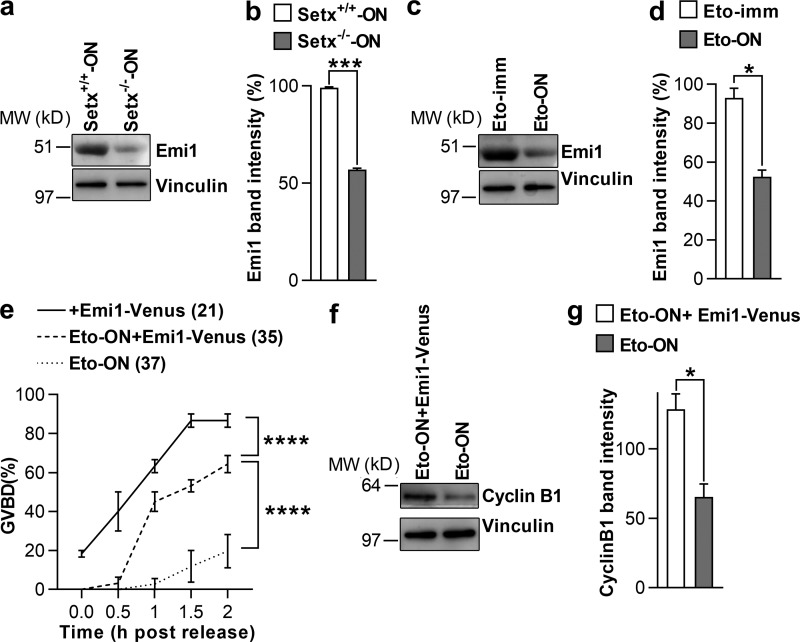
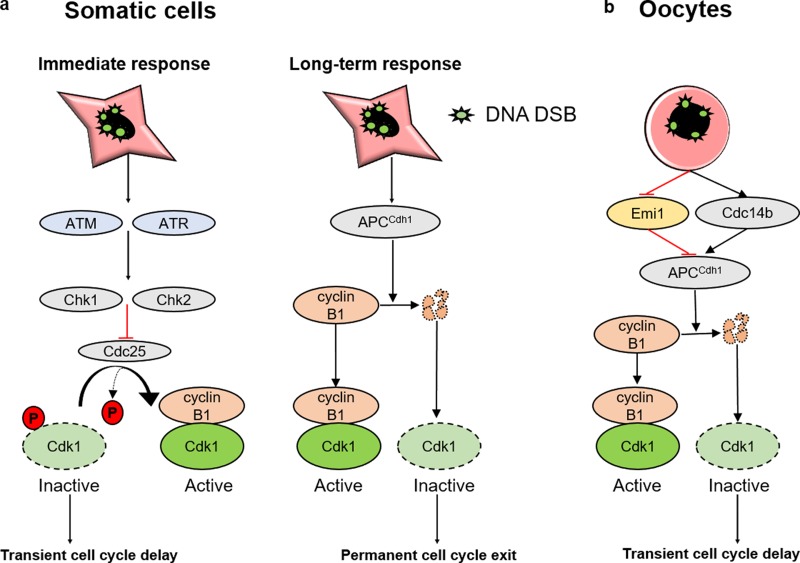
In mitotic cells, DNA damage induces temporary G2 arrest via inhibitory Cdk1 phosphorylation. In contrast, fully grown G2-stage oocytes readily enter M phase immediately following chemical induction of DNA damage in vitro, indicating that the canonical immediate-response G2/M DNA damage response (DDR) may be deficient. Senataxin (Setx) is involved in RNA/DNA processing and maintaining genome integrity. Here we find that mouse oocytes deleted of Setx accumulate DNA damage when exposed to oxidative stress in vitro and during aging in vivo, after which, surprisingly, they undergo G2 arrest. Moreover, fully grown wild-type oocytes undergo G2 arrest after chemotherapy-induced in vitro damage if an overnight delay is imposed following damage induction. Unexpectedly, this slow-evolving DDR is not mediated by inhibitory Cdk1 phosphorylation but by APC-Cdh1-mediated proteolysis of the Cdk1 activator, cyclin B1, secondary to increased Cdc14B-dependent APC-Cdh1 activation and reduced Emi1-dependent inhibition. Thus, oocytes are unable to respond immediately to DNA damage, but instead mount a G2/M DDR that evolves slowly and involves a phosphorylation-independent proteolytic pathway.
© 2020 Subramanian et al.
Figures
References
-
- Adhikari D., Busayavalasa K., Zhang J., Hu M., Risal S., Bayazit M.B., Singh M., Diril M.K., Kaldis P., and Liu K.. 2016. Inhibitory phosphorylation of Cdk1 mediates prolonged prophase I arrest in female germ cells and is essential for female reproductive lifespan. Cell Res. 26:1212–1225. 10.1038/cr.2016.119 - DOI - PMC - PubMed
-
- Becherel O.J., Yeo A.J., Stellati A., Heng E.Y., Luff J., Suraweera A.M., Woods R., Fleming J., Carrie D., McKinney K., et al. 2013. Senataxin plays an essential role with DNA damage response proteins in meiotic recombination and gene silencing. PLoS Genet. 9:e1003435 10.1371/journal.pgen.1003435 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
