

TRIC-A shapes oscillatory Ca2+ signals by interaction with STIM1/Orai1 complexes
- PMID: 32330125
- PMCID: PMC7202670
- DOI: 10.1371/journal.pbio.3000700
TRIC-A shapes oscillatory Ca2+ signals by interaction with STIM1/Orai1 complexes
Abstract
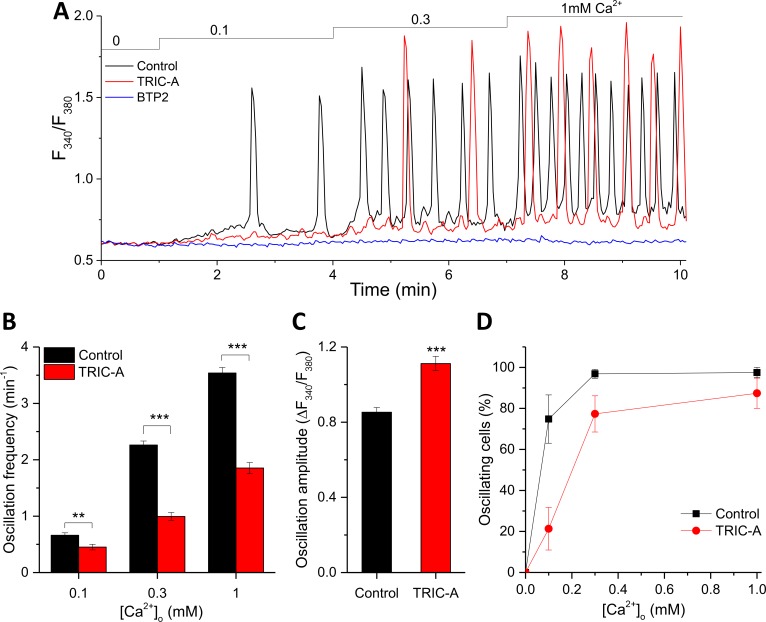
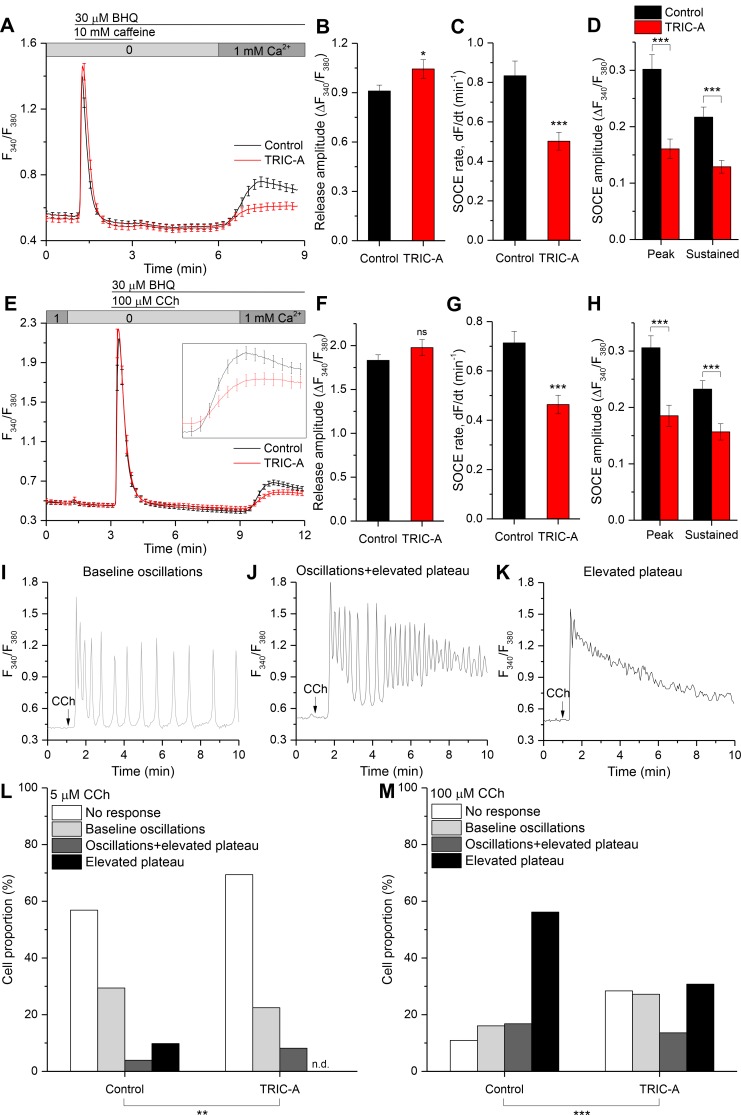
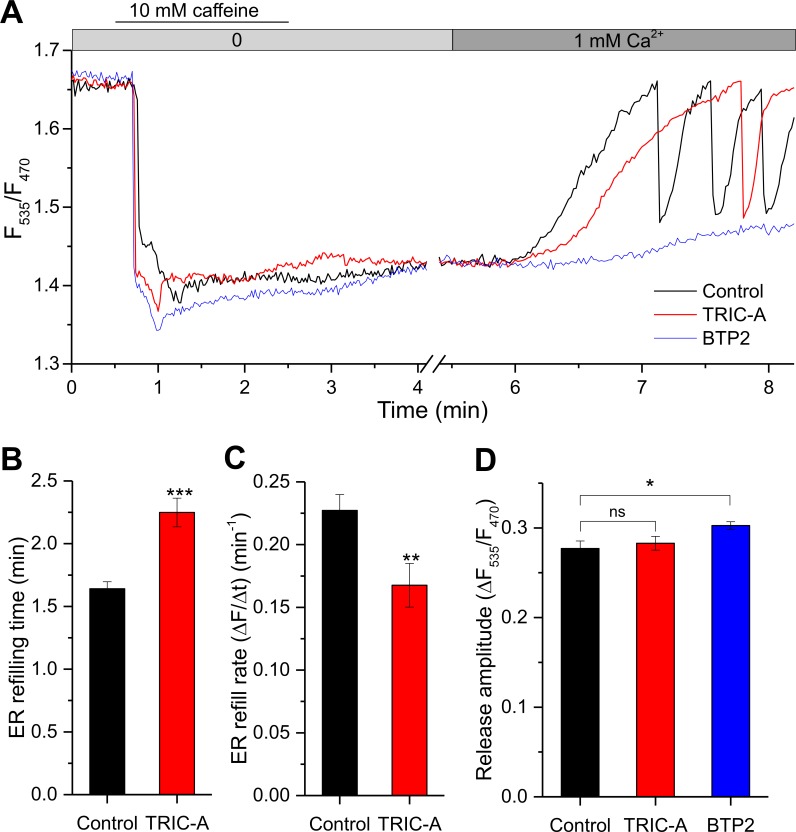
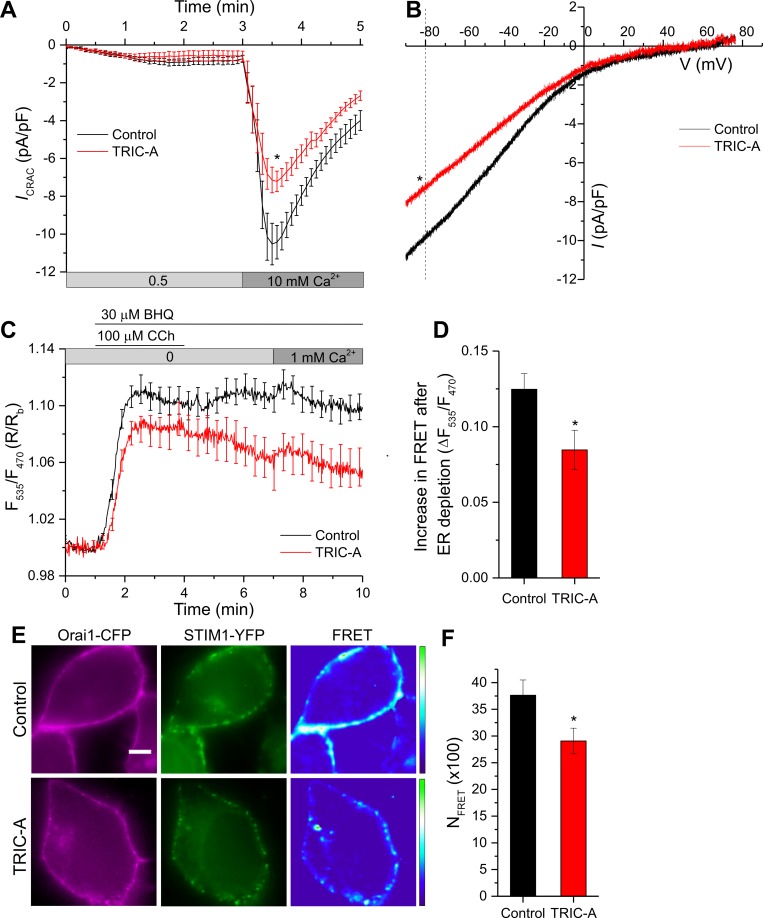
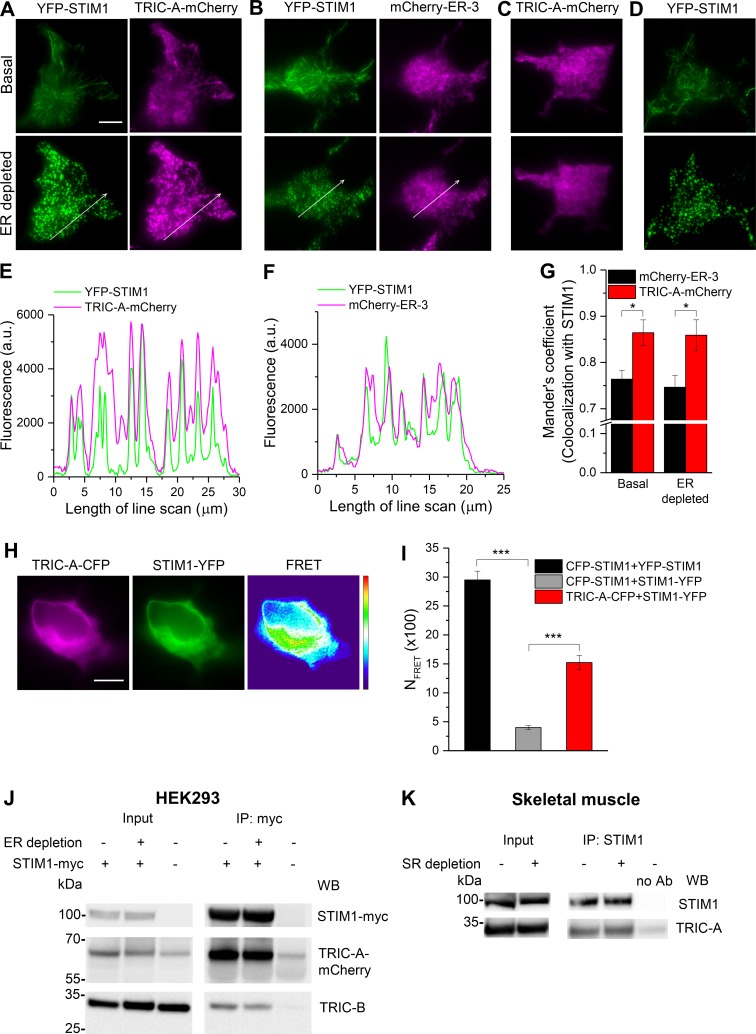
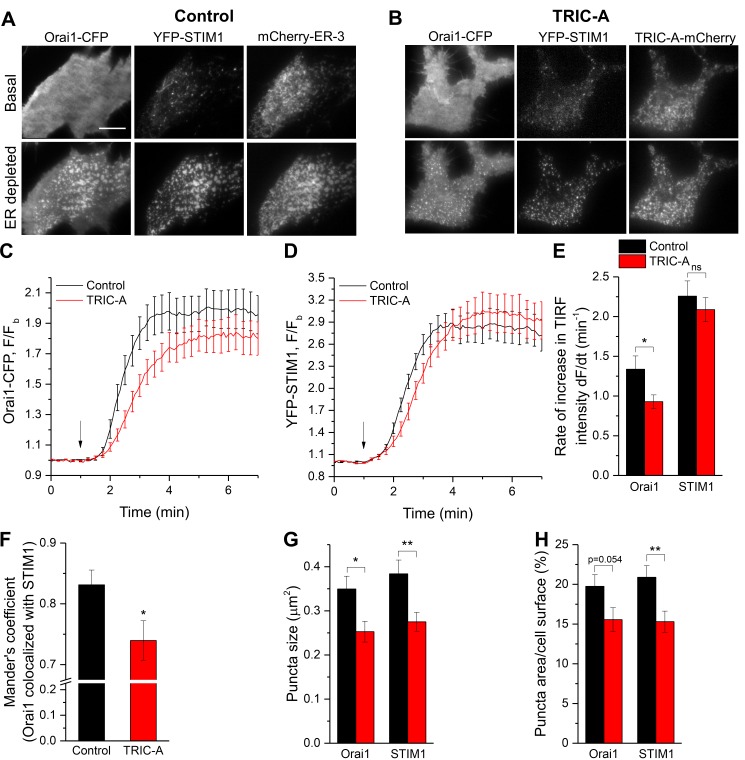
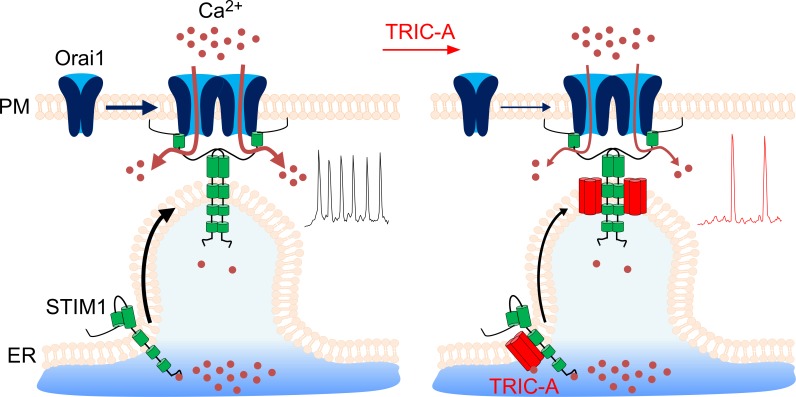
Trimeric intracellular cation (TRIC) channels have been proposed to modulate Ca2+ release from the endoplasmic reticulum (ER) and determine oscillatory Ca2+ signals. Here, we report that TRIC-A-mediated amplitude and frequency modulation of ryanodine receptor 2 (RyR2)-mediated Ca2+ oscillations and inositol 1,4,5-triphosphate receptor (IP3R)-induced cytosolic signals is based on attenuating store-operated Ca2+ entry (SOCE). Further, TRIC-A-dependent delay in ER Ca2+ store refilling contributes to shaping the pattern of Ca2+ oscillations. Upon ER Ca2+ depletion, TRIC-A clusters with stromal interaction molecule 1 (STIM1) and Ca2+-release-activated Ca2+ channel 1 (Orai1) within ER-plasma membrane (PM) junctions and impairs assembly of the STIM1/Orai1 complex, causing a decrease in Orai1-mediated Ca2+ current and SOCE. Together, our findings demonstrate that TRIC-A is a negative regulator of STIM1/Orai1 function. Thus, aberrant SOCE could contribute to muscle disorders associated with loss of TRIC-A.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Comment in
-
TRICking SOCE into altered oscillations.Cell Calcium. 2020 Dec;92:102290. doi: 10.1016/j.ceca.2020.102290. Epub 2020 Sep 22. Cell Calcium. 2020. PMID: 32979765
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
