

Engineering the Live-Attenuated Polio Vaccine to Prevent Reversion to Virulence
- PMID: 32330425
- PMCID: PMC7566161
- DOI: 10.1016/j.chom.2020.04.003
Engineering the Live-Attenuated Polio Vaccine to Prevent Reversion to Virulence
Abstract
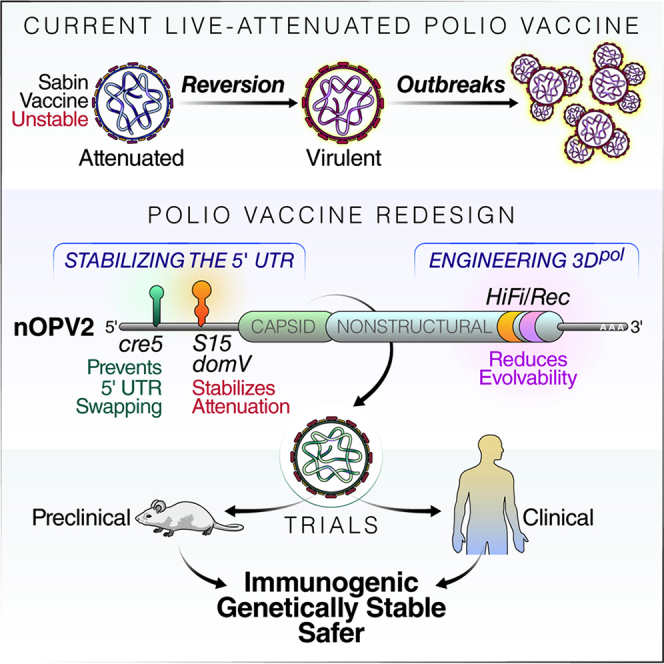
The live-attenuated oral poliovirus vaccine (OPV or Sabin vaccine) replicates in gut-associated tissues, eliciting mucosa and systemic immunity. OPV protects from disease and limits poliovirus spread. Accordingly, vaccination with OPV is the primary strategy used to end the circulation of all polioviruses. However, the ability of OPV to regain replication fitness and establish new epidemics represents a significant risk of polio re-emergence should immunization cease. Here, we report the development of a poliovirus type 2 vaccine strain (nOPV2) that is genetically more stable and less likely to regain virulence than the original Sabin2 strain. We introduced modifications within at the 5' untranslated region of the Sabin2 genome to stabilize attenuation determinants, 2C coding region to prevent recombination, and 3D polymerase to limit viral adaptability. Prior work established that nOPV2 is immunogenic in preclinical and clinical studies, and thus may enable complete poliovirus eradication.
Keywords: OPV2; Sabin 2p; live-attenuated vaccine; poliovirus; poliovirus eradication.
Copyright © 2020. Published by Elsevier Inc.
Conflict of interest statement
Declaration of Interests The authors declare no competing interests.
Figures
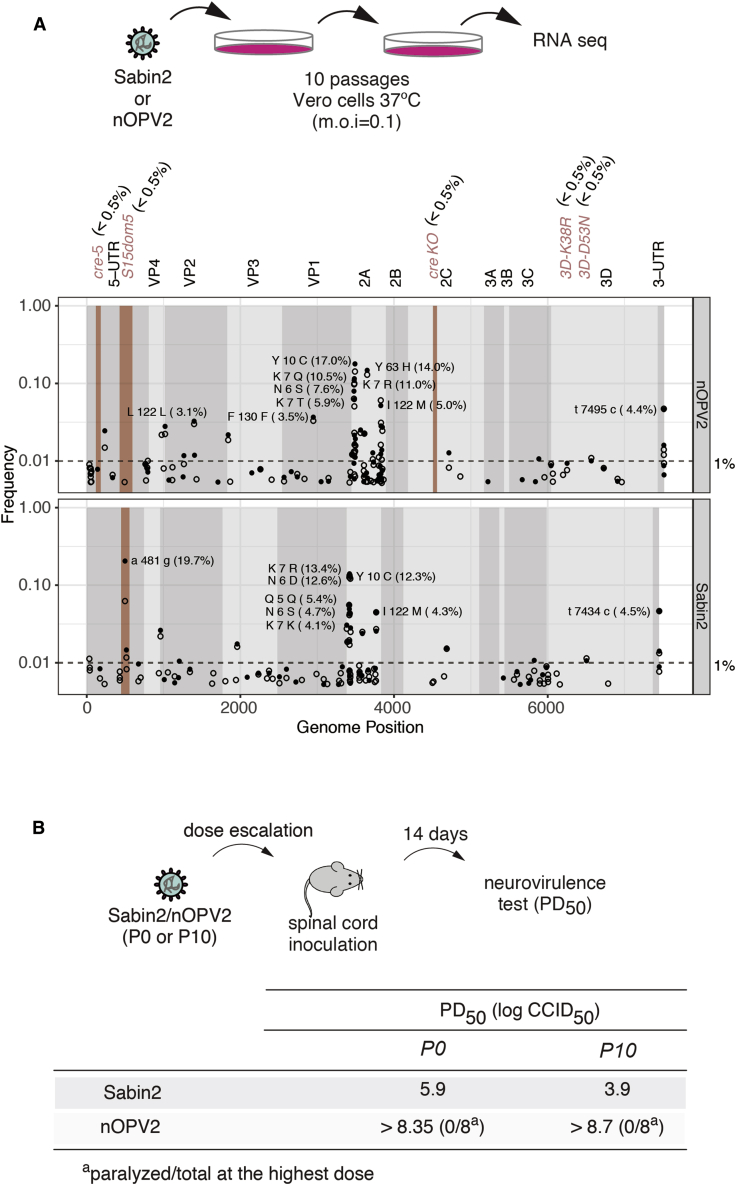
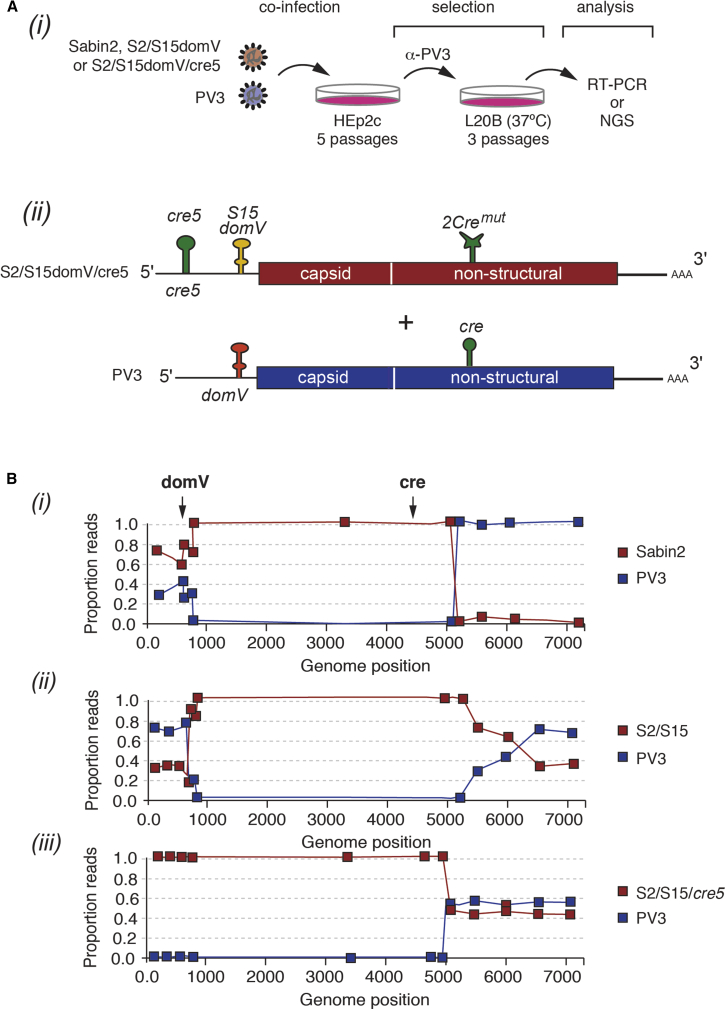
Comment in
-
New polio vaccine poised to get emergency WHO approval.Nature. 2020 Nov;587(7832):15-16. doi: 10.1038/d41586-020-03045-2. Nature. 2020. PMID: 33122836 No abstract available.
References
-
- Asturias E.J., Bandyopadhyay A.S., Self S., Rivera L., Sáez-Llorens X., Lopez E., Melgar M., Gaensbauer J.T., Weldon W.C., Oberste M.S., Latin American IPV001BMG Study Group Humoral and intestinal immunity induced by new schedules of bivalent oral poliovirus vaccine and one or two doses of inactivated poliovirus vaccine in Latin American infants: an open-label randomised controlled trial. Lancet. 2016;388:158–169. - PubMed
-
- Bandyopadhyay A.S., Garon J., Seib K., Orenstein W.A. Polio vaccination: past, present and future. Future Microbiol. 2015;10:791–808. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
