

Palmitoylation of the KATP channel Kir6.2 subunit promotes channel opening by regulating PIP2 sensitivity
- PMID: 32332165
- PMCID: PMC7229695
- DOI: 10.1073/pnas.1918088117
Palmitoylation of the KATP channel Kir6.2 subunit promotes channel opening by regulating PIP2 sensitivity
Abstract
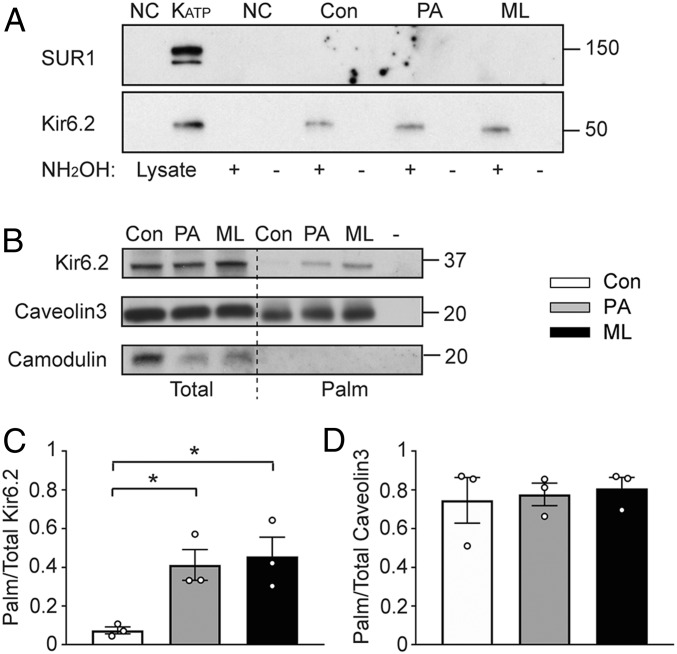
A physiological role for long-chain acyl-CoA esters to activate ATP-sensitive K+ (KATP) channels is well established. Circulating palmitate is transported into cells and converted to palmitoyl-CoA, which is a substrate for palmitoylation. We found that palmitoyl-CoA, but not palmitic acid, activated the channel when applied acutely. We have altered the palmitoylation state by preincubating cells with micromolar concentrations of palmitic acid or by inhibiting protein thioesterases. With acyl-biotin exchange assays we found that Kir6.2, but not sulfonylurea receptor (SUR)1 or SUR2, was palmitoylated. These interventions increased the KATP channel mean patch current, increased the open time, and decreased the apparent sensitivity to ATP without affecting surface expression. Similar data were obtained in transfected cells, rat insulin-secreting INS-1 cells, and isolated cardiac myocytes. Kir6.2ΔC36, expressed without SUR, was also positively regulated by palmitoylation. Mutagenesis of Kir6.2 Cys166 prevented these effects. Clinical variants in KCNJ11 that affect Cys166 had a similar gain-of-function phenotype, but was more pronounced. Molecular modeling studies suggested that palmitoyl-C166 and selected large hydrophobic mutations make direct hydrophobic contact with Kir6.2-bound PIP2 Patch-clamp studies confirmed that palmitoylation of Kir6.2 at Cys166 enhanced the PIP2 sensitivity of the channel. Physiological relevance is suggested since palmitoylation blunted the regulation of KATP channels by α1-adrenoreceptor stimulation. The Cys166 residue is conserved in some other Kir family members (Kir6.1 and Kir3, but not Kir2), which are also subject to regulated palmitoylation, suggesting a general mechanism to control the open state of certain Kir channels.
Keywords: KATP channels; Kir6.2; PIP2; lipidation; palmitoylation.
Conflict of interest statement
The authors declare no competing interest.
Figures







References
-
- Lin D. T., Conibear E., Enzymatic protein depalmitoylation by acyl protein thioesterases. Biochem. Soc. Trans. 43, 193–198 (2015). - PubMed
-
- Aicart-Ramos C., Valero R. A., Rodriguez-Crespo I., Protein palmitoylation and subcellular trafficking. Biochim. Biophys. Acta 1808, 2981–2994 (2011). - PubMed
-
- Tortosa E., Hoogenraad C. C., Polarized trafficking: The palmitoylation cycle distributes cytoplasmic proteins to distinct neuronal compartments. Curr. Opin. Cell Biol. 50, 64–71 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
