

Decreased brain and muscle ARNT-like protein 1 expression mediated the contribution of hyperandrogenism to insulin resistance in polycystic ovary syndrome
- PMID: 32334629
- PMCID: PMC7183135
- DOI: 10.1186/s12958-020-00592-1
Decreased brain and muscle ARNT-like protein 1 expression mediated the contribution of hyperandrogenism to insulin resistance in polycystic ovary syndrome
Abstract
Background: The interface between environmental risk factors and genetic factors could contribute to the pathogenesis of hyperandrogenism and insulin resistance in polycystic ovary syndrome (PCOS); however, the underlying complex mechanism remains to be elucidated.
Methods: We used dehydroepiandrosterone (DHEA)-induced PCOS-like rat model to measure circadian clock genes and insulin resistance-related genes. Additionally, we performed in vitro experiments in mature adipocytes to verify the molecular mechanisms.
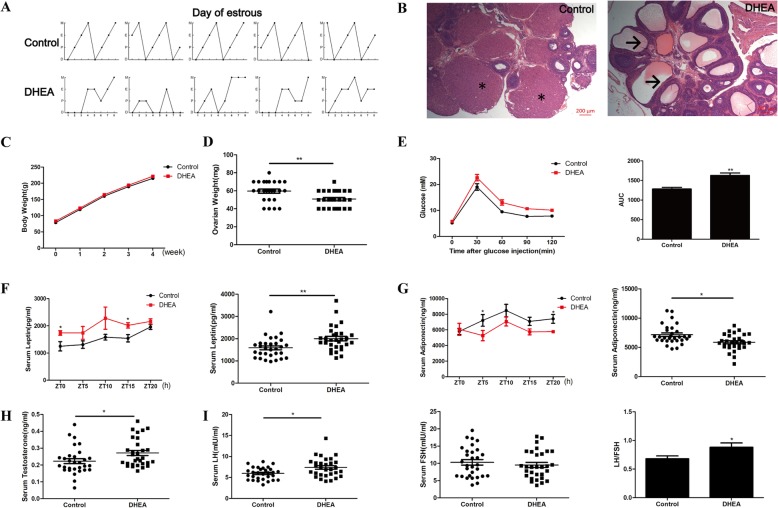
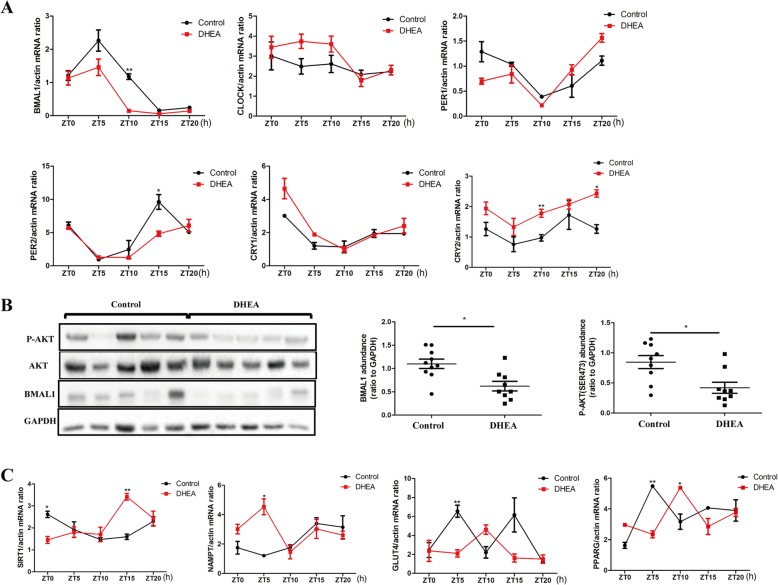
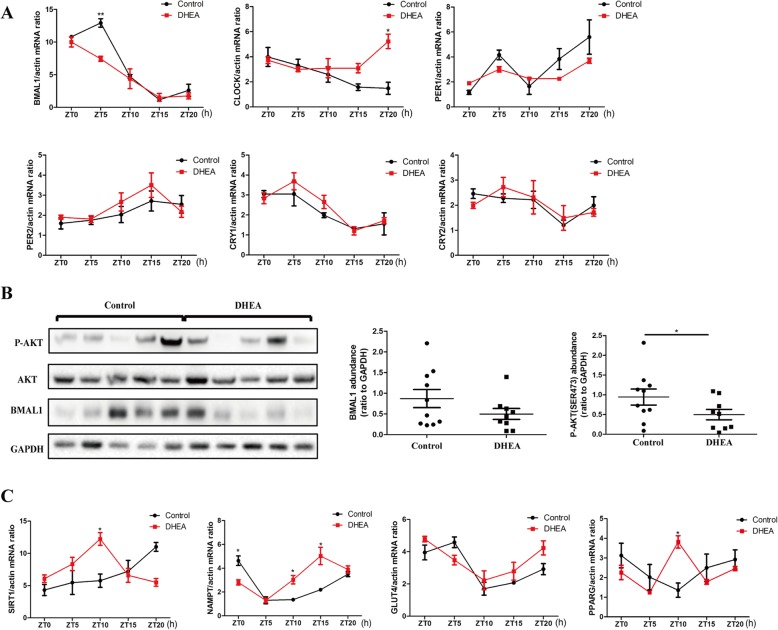
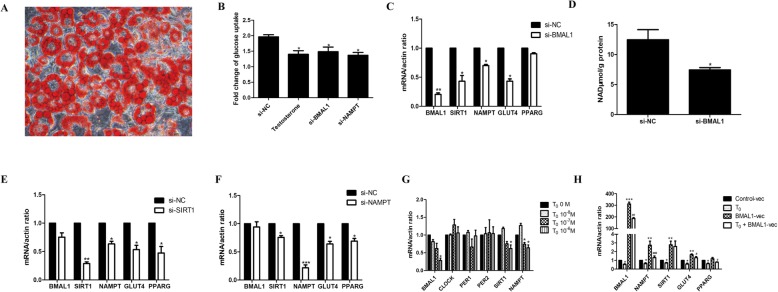
Results: DHEA-induced PCOS-like rats exhibited insulin resistance and arrhythmic expression of circadian clock genes in the liver and adipose tissues, particularly showing decreased brain and muscle ARNT-like protein 1 (BMAL1) expression. In addition, hyperandrogenism gave rise to negative regulation of BMAL1 expression to nicotinamide phosphoribosyltransferase and sirtuin 1, which further inhibited downstream glucose transporter type 4, leading to insulin resistance in mature adipocytes, which was consistent with our previous results in HepG2 cells.
Conclusions: Decreased BMAL1 expression in the liver and adipose played a potentially novel role in the contribution of hyperandrogenism to insulin resistance, which might be a possible mechanism accounting for the pathogenesis of PCOS.
Keywords: BMAL1; Circadian clock; Hyperandrogenism; Insulin resistance; PCOS.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
References
MeSH terms
Substances
Grants and funding
- 2018YFC1003202/National Key Research and Development Program of China
- 2017YFC1001002/National Key Research and Development Program of China
- 81971343/National Natural Science Foundation of China
- 81901549/National Natural Science Foundation of China
- 81671414/National Natural Science Foundation of China
LinkOut - more resources
Full Text Sources
Medical
Research Materials
