

Mutations in SPATA13/ASEF2 cause primary angle closure glaucoma
- PMID: 32339198
- PMCID: PMC7233598
- DOI: 10.1371/journal.pgen.1008721
Mutations in SPATA13/ASEF2 cause primary angle closure glaucoma
Abstract
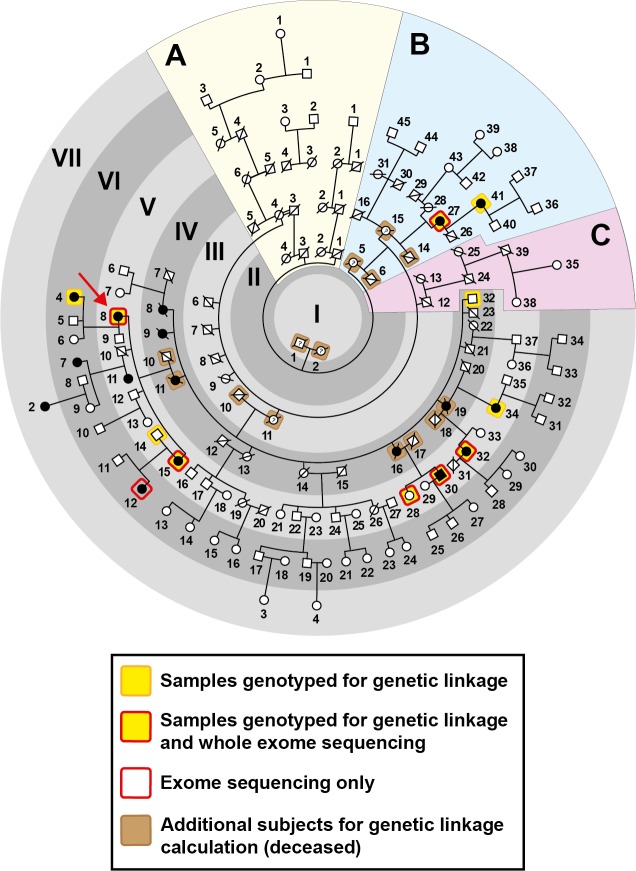
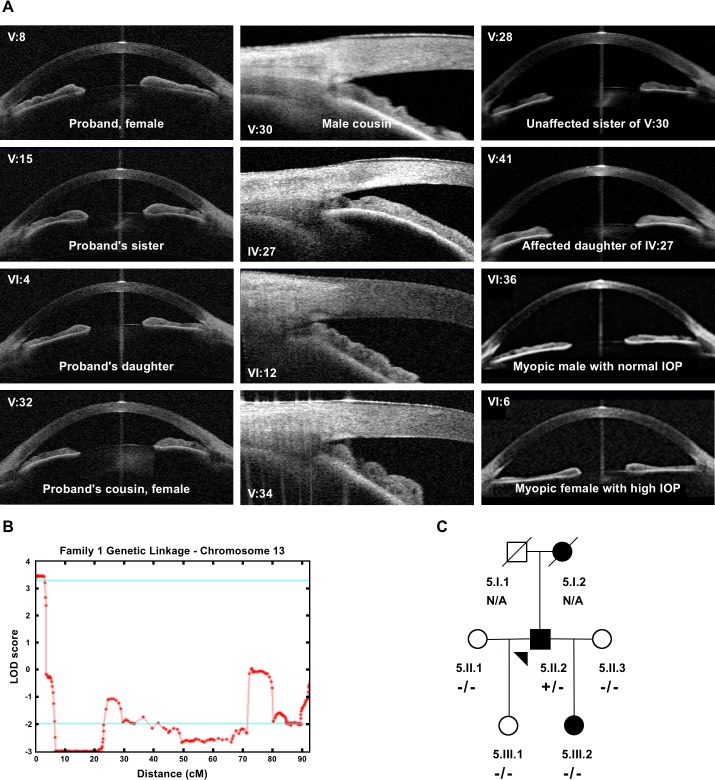
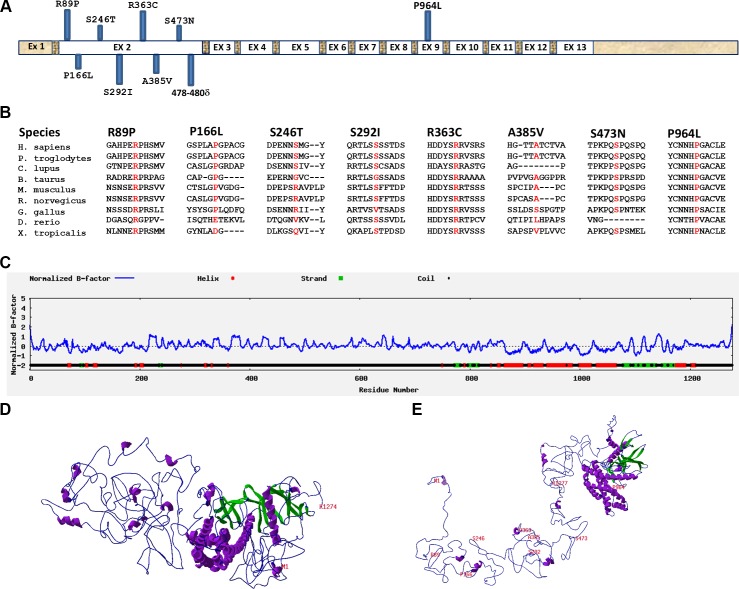
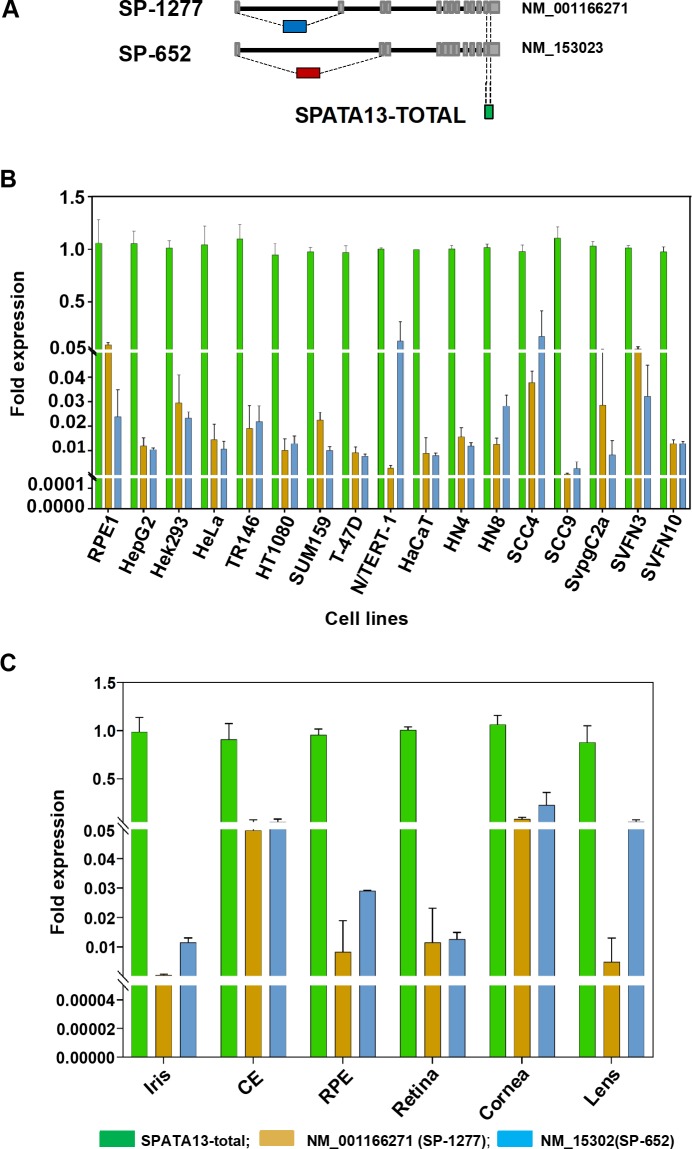
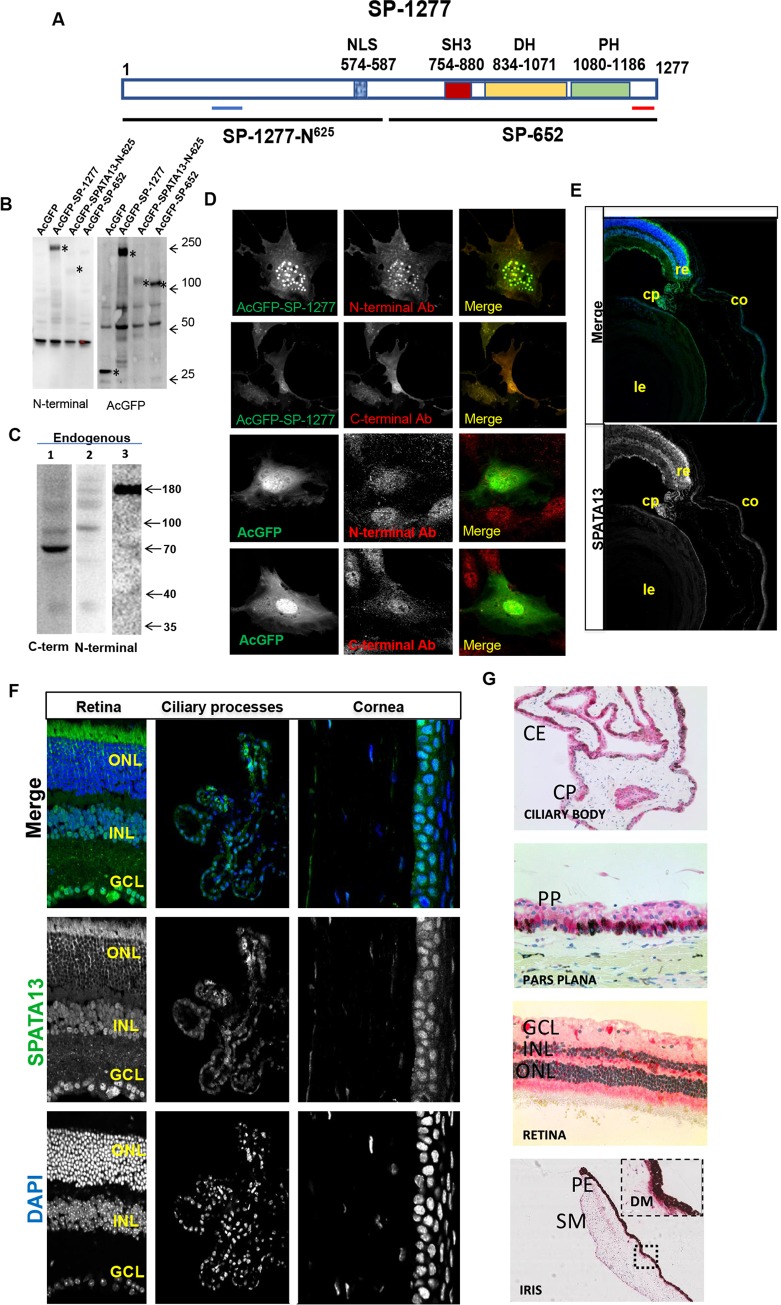
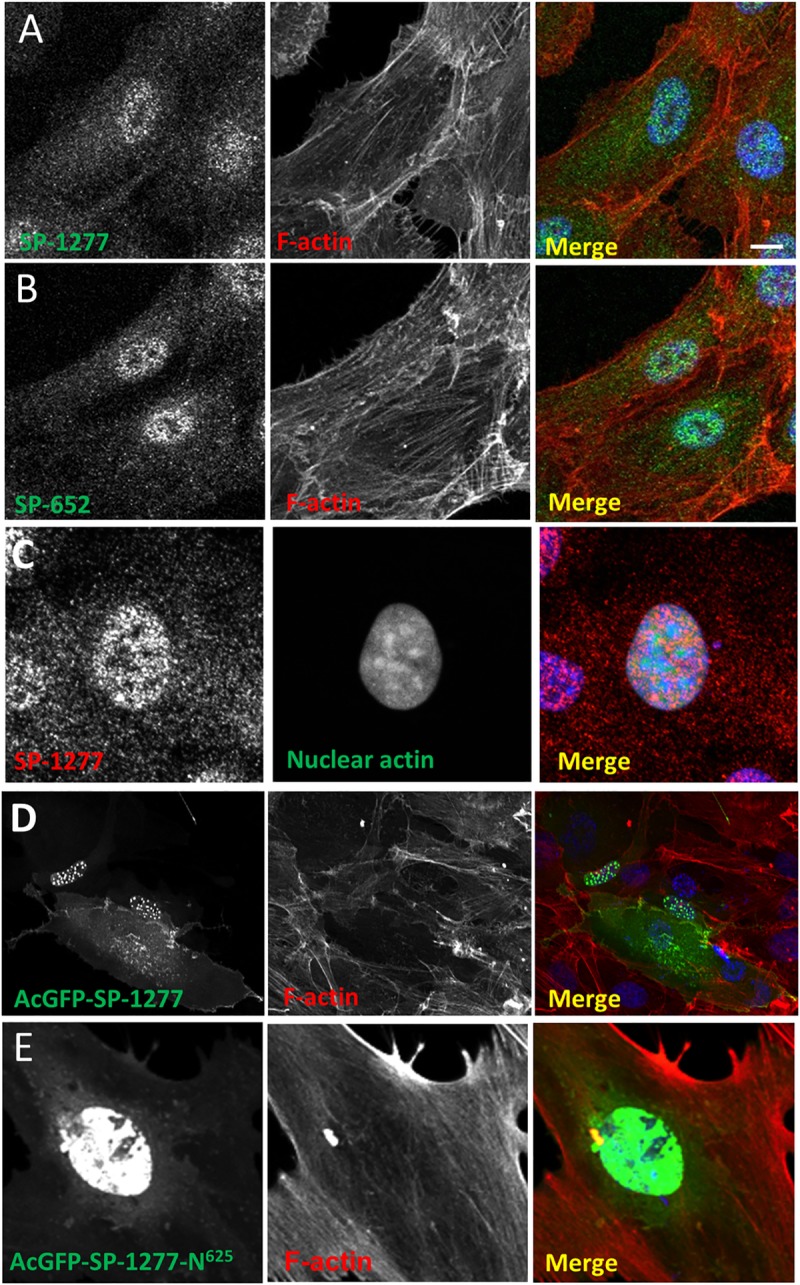
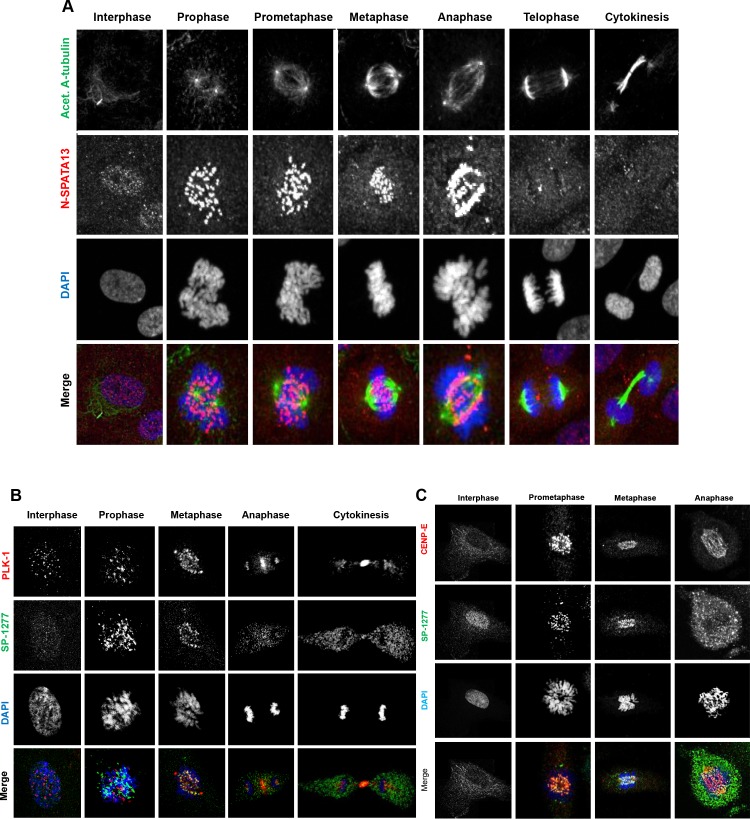
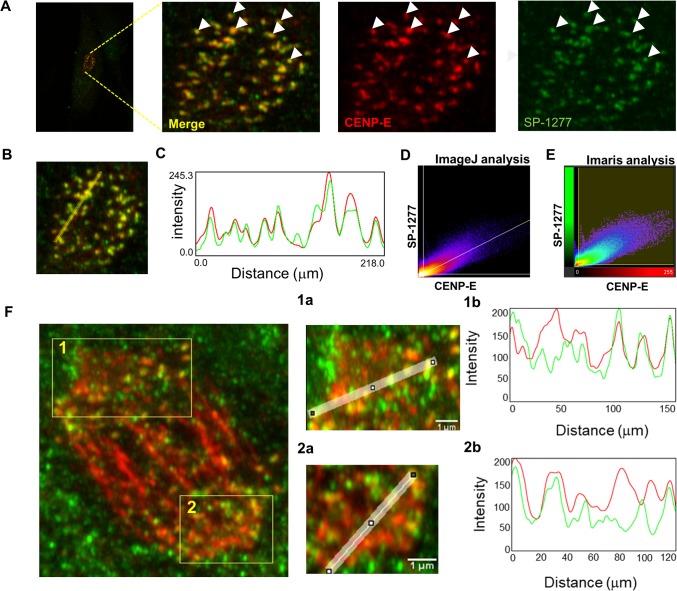
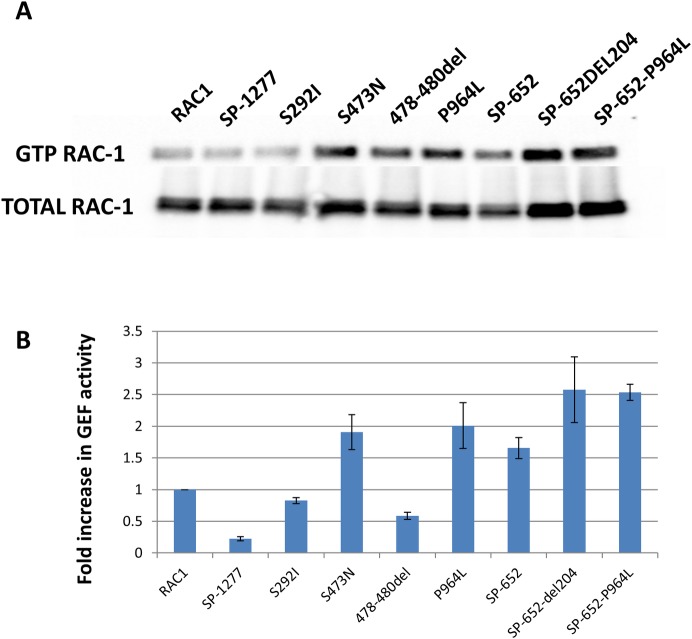
Current estimates suggest 50% of glaucoma blindness worldwide is caused by primary angle-closure glaucoma (PACG) but the causative gene is not known. We used genetic linkage and whole genome sequencing to identify Spermatogenesis Associated Protein 13, SPATA13 (NM_001166271; NP_001159743, SPATA13 isoform I), also known as ASEF2 (Adenomatous polyposis coli-stimulated guanine nucleotide exchange factor 2), as the causal gene for PACG in a large seven-generation white British family showing variable expression and incomplete penetrance. The 9 bp deletion, c.1432_1440del; p.478_480del was present in all affected individuals with angle-closure disease. We show ubiquitous expression of this transcript in cell lines derived from human tissues and in iris, retina, retinal pigment and ciliary epithelia, cornea and lens. We also identified eight additional mutations in SPATA13 in a cohort of 189 unrelated PACS/PAC/PACG samples. This gene encodes a 1277 residue protein which localises to the nucleus with partial co-localisation with nuclear speckles. In cells undergoing mitosis SPATA13 isoform I becomes part of the kinetochore complex co-localising with two kinetochore markers, polo like kinase 1 (PLK-1) and centrosome-associated protein E (CENP-E). The 9 bp deletion reported in this study increases the RAC1-dependent guanine nucleotide exchange factors (GEF) activity. The increase in GEF activity was also observed in three other variants identified in this study. Taken together, our data suggest that SPATA13 is involved in the regulation of mitosis and the mutations dysregulate GEF activity affecting homeostasis in tissues where it is highly expressed, influencing PACG pathogenesis.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
