

Plant NLR immune receptor Tm-22 activation requires NB-ARC domain-mediated self-association of CC domain
- PMID: 32339200
- PMCID: PMC7205312
- DOI: 10.1371/journal.ppat.1008475
Plant NLR immune receptor Tm-22 activation requires NB-ARC domain-mediated self-association of CC domain
Abstract
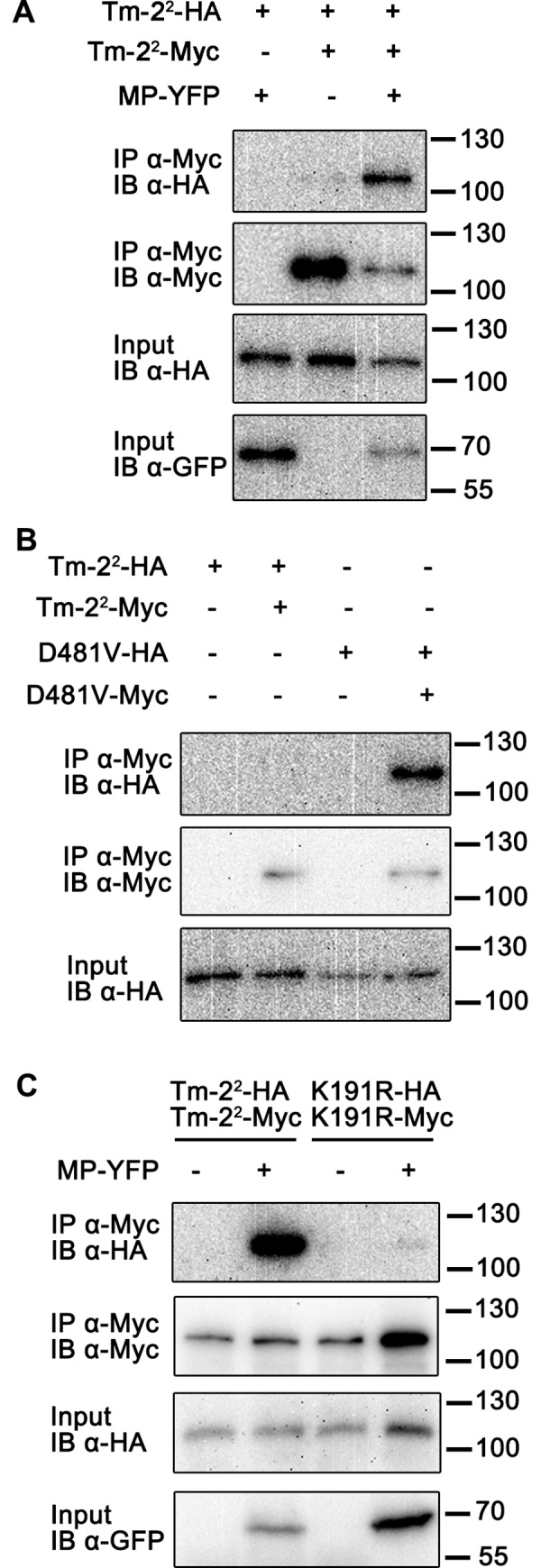
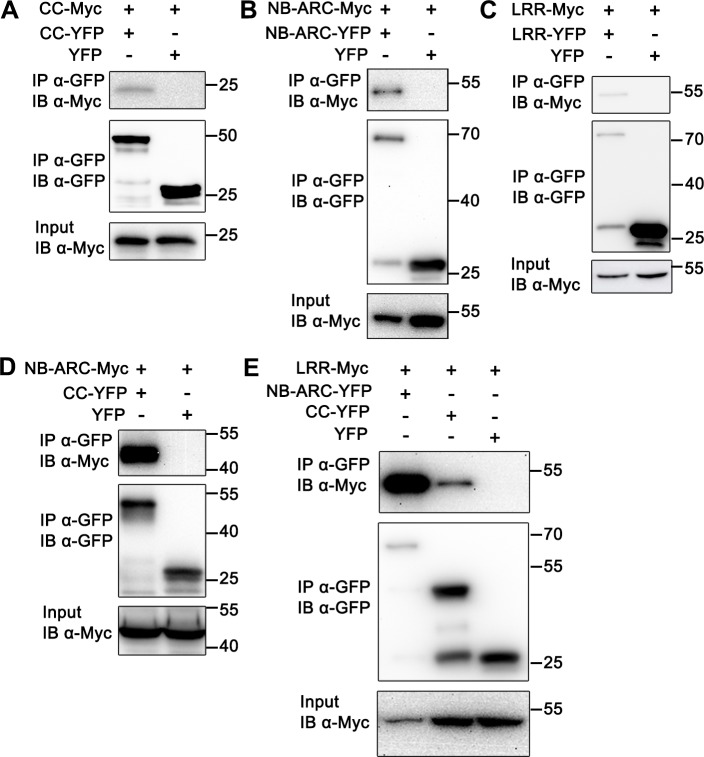
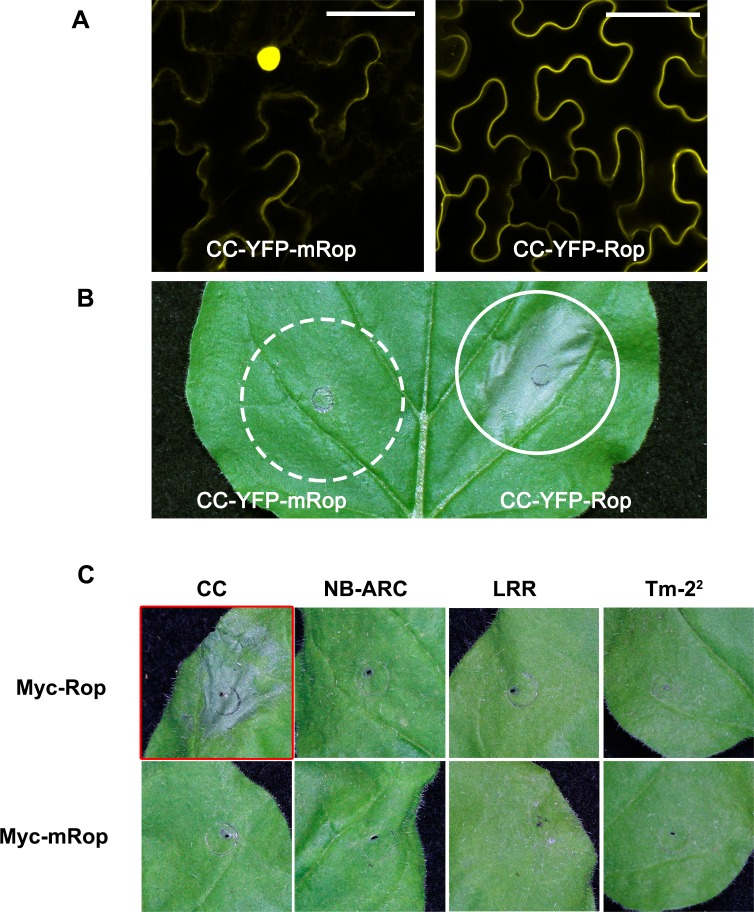
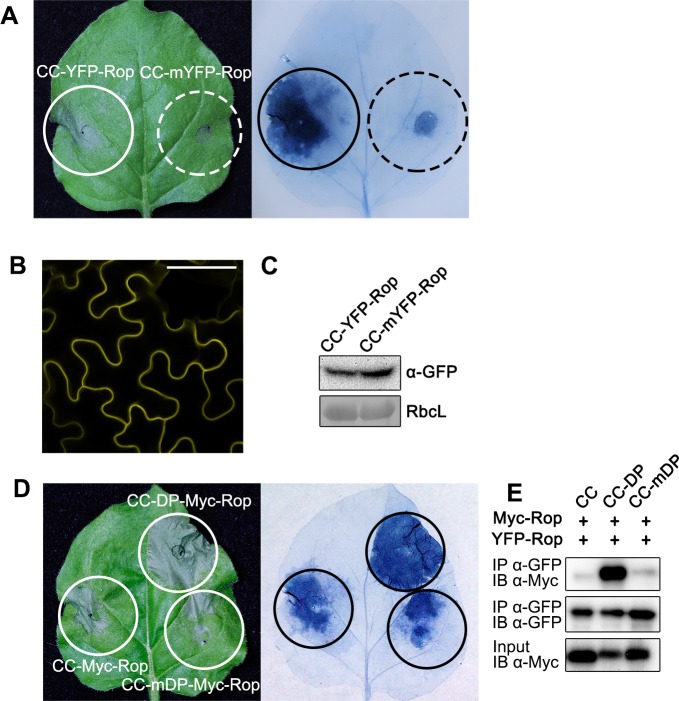
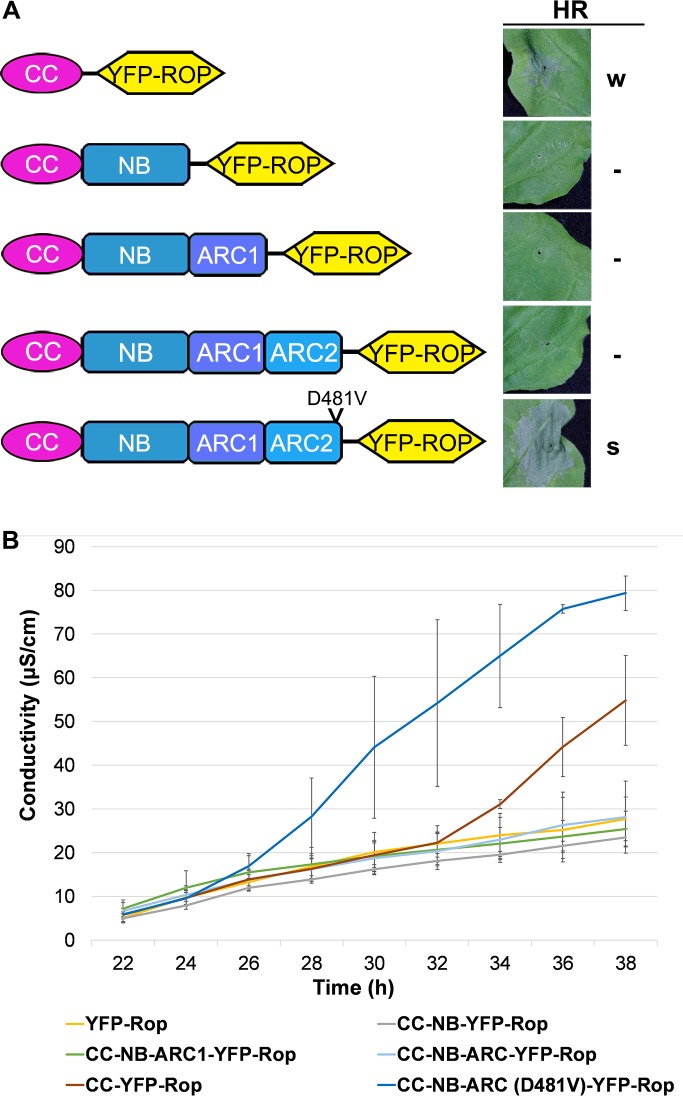
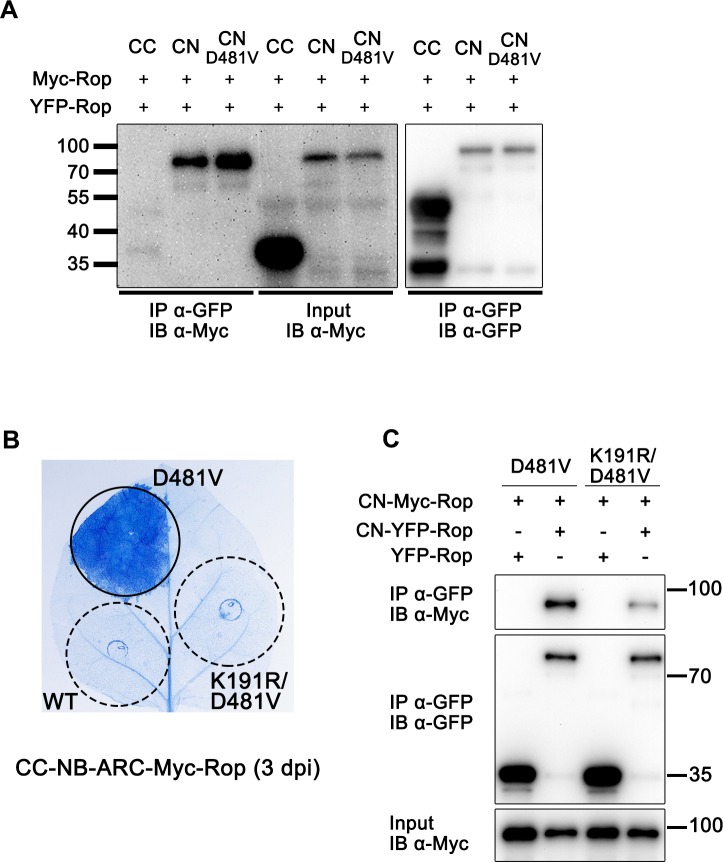
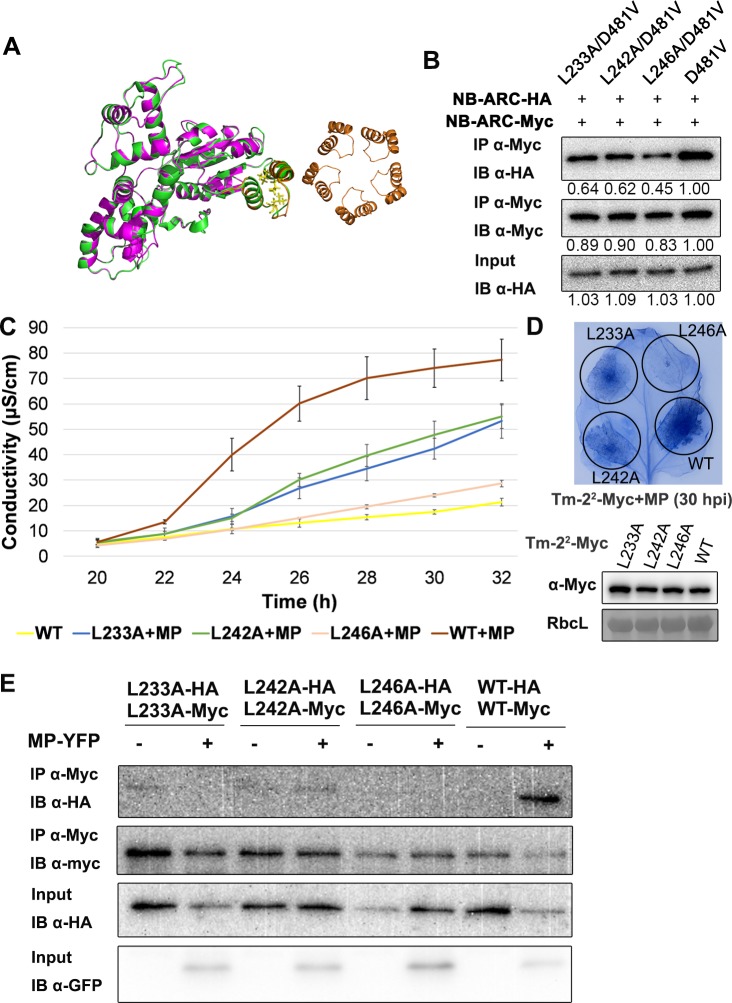
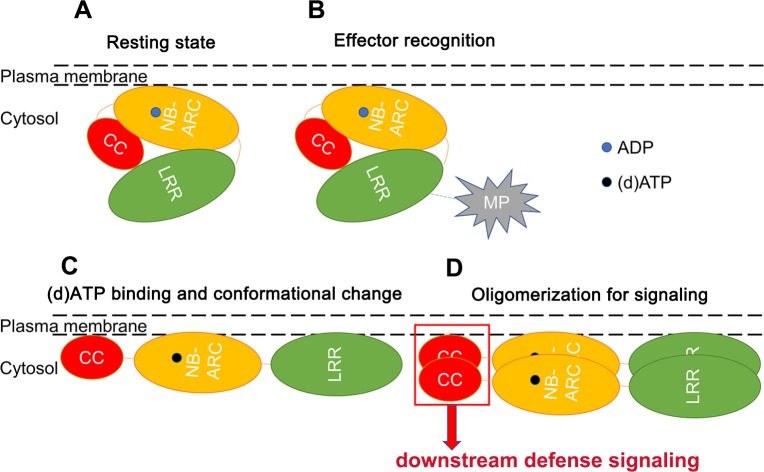
The nucleotide-binding, leucine-rich repeat-containing (NLR) class of immune receptors of plants and animals recognize pathogen-encoded proteins and trigger host defenses. Although animal NLRs form oligomers upon pathogen recognition to activate downstream signaling, the mechanisms of plant NLR activation remain largely elusive. Tm-22 is a plasma membrane (PM)-localized coiled coil (CC)-type NLR and confers resistance to Tobacco mosaic virus (TMV) by recognizing its viral movement protein (MP). In this study, we found that Tm-22 self-associates upon recognition of MP. The CC domain of Tm-22 is the signaling domain and its function requires PM localization and self-association. The nucleotide-binding (NB-ARC) domain is important for Tm-22 self-interaction and regulates activation of the CC domain through its nucleotide-binding and self-association. (d)ATP binding may alter the NB-ARC conformation to release its suppression of Tm-22 CC domain-mediated cell death. Our findings provide the first example of signaling domain for PM-localized NLR and insight into PM-localized NLR activation.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Jones JD, Vance RE, Dangl JL. Intracellular innate immune surveillance devices in plants and animals. Science. 2016;354(6316). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
