

Vaccines and Broadly Neutralizing Antibodies for HIV-1 Prevention
- PMID: 32340576
- PMCID: PMC7375352
- DOI: 10.1146/annurev-immunol-080219-023629
Vaccines and Broadly Neutralizing Antibodies for HIV-1 Prevention
Abstract
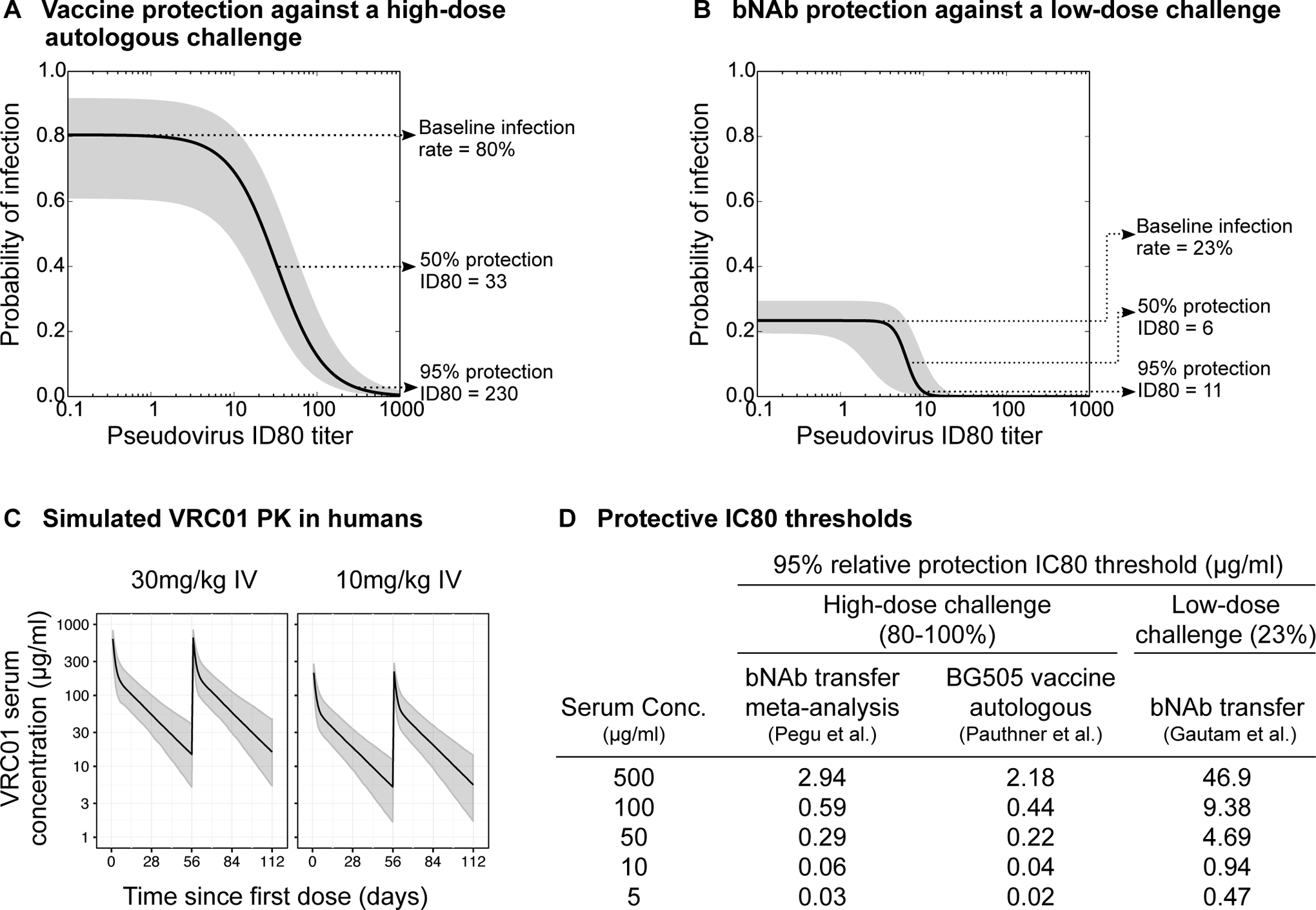
Development of improved approaches for HIV-1 prevention will likely be required for a durable end to the global AIDS pandemic. Recent advances in preclinical studies and early phase clinical trials offer renewed promise for immunologic strategies for blocking acquisition of HIV-1 infection. Clinical trials are currently underway to evaluate the efficacy of two vaccine candidates and a broadly neutralizing antibody (bNAb) to prevent HIV-1 infection in humans. However, the vast diversity of HIV-1 is a major challenge for both active and passive immunization. Here we review current immunologic strategies for HIV-1 prevention, with a focus on current and next-generation vaccines and bNAbs.
Keywords: HIV-1; bNAb; broadly neutralizing antibody; prevention; vaccine; viral diversity.
Figures





References
-
- UNAIDS. 2018. UNAIDS Data 2018.
-
- UNAIDS. 2015. UNAIDS 2016–2021 Strategy: On the Fast-Track to end AIDS
-
- Weller S, Davis K. 2002. Condom effectiveness in reducing heterosexual HIV transmission. The Cochrane Database of Systematic Reviews: CD003255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
