

Structure and Function of Influenza Polymerase
- PMID: 32341065
- PMCID: PMC8415296
- DOI: 10.1101/cshperspect.a038372
Structure and Function of Influenza Polymerase
Abstract
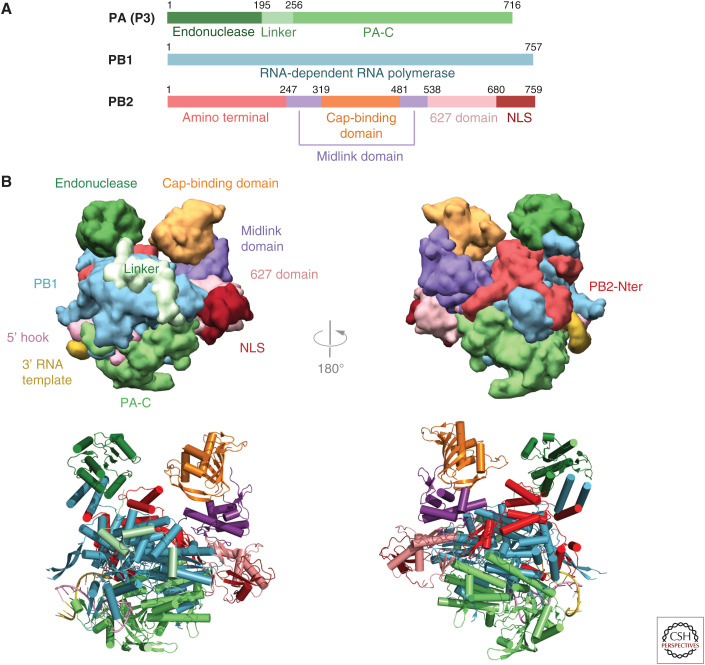
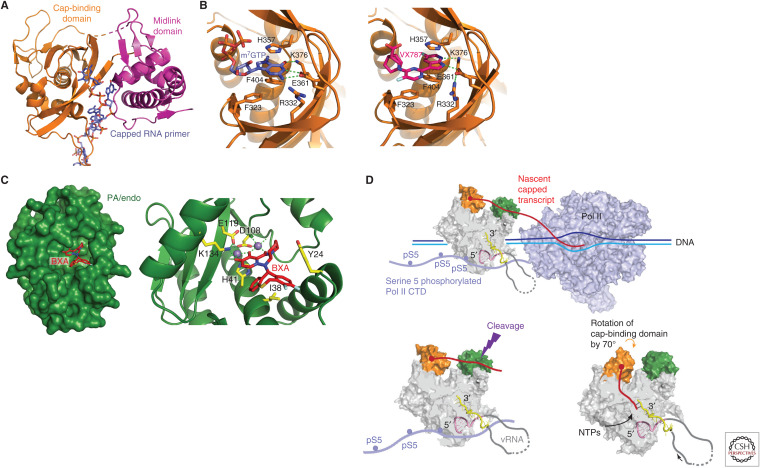
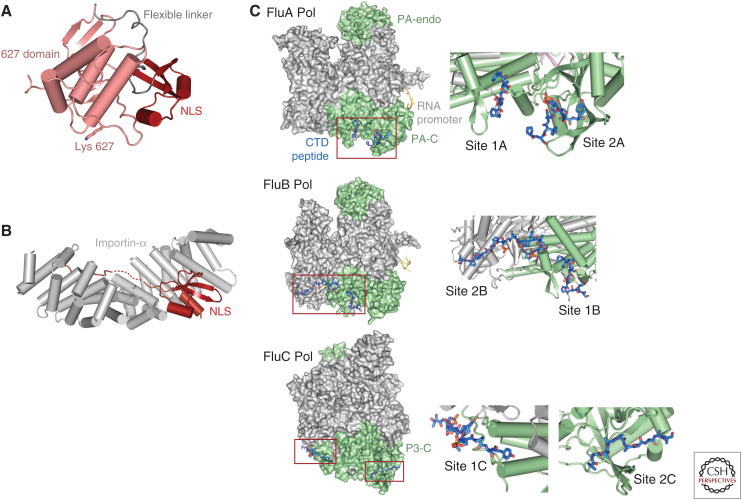
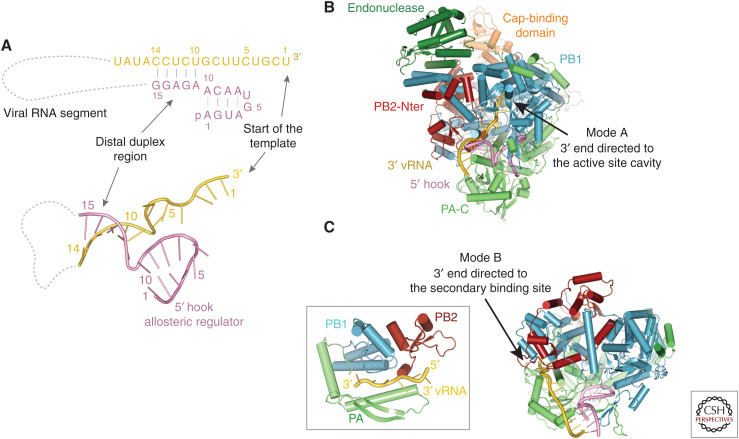
Influenza polymerase (FluPol) plays a key role in the viral infection cycle by transcribing and replicating the viral genome. FluPol is a multifunctional, heterotrimeric enzyme with cap-binding, endonuclease, RNA-dependent RNA polymerase and polyadenylation activities. It performs its functions in the context of the viral ribonucleoprotein particle (RNP), wherein the template viral RNA is coated by multiple copies of viral nucleoprotein. Moreover, it interacts with a number of host proteins that are essential cofactors and, consequently, adaptive mutations in the polymerase are required for crossing the avian-human species barrier. In this review, we show how mechanistic understanding of how FluPol performs its multiple functions has greatly advanced over the last decade through determination of high-resolution structures by X-ray crystallography and cryo-electron microscopy. These have revealed not only the detailed architecture of FluPol but highlighted the remarkably conformational flexibility that is inherent to its functioning as a dynamic RNA synthesis machine. Structural studies are also underpinning current attempts to develop next-generation anti-influenza drugs that directly target FluPol.
Copyright © 2021 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures

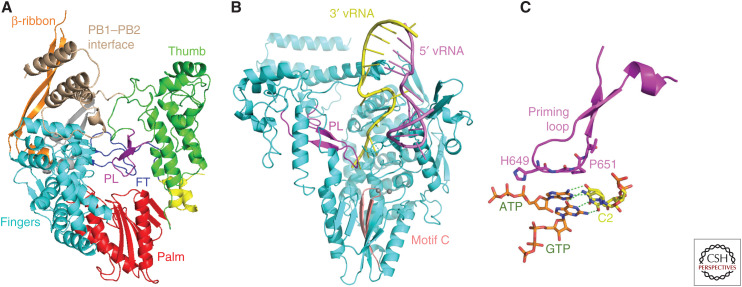
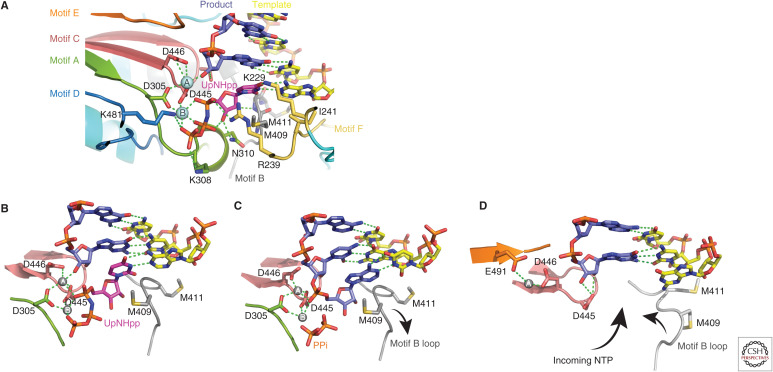
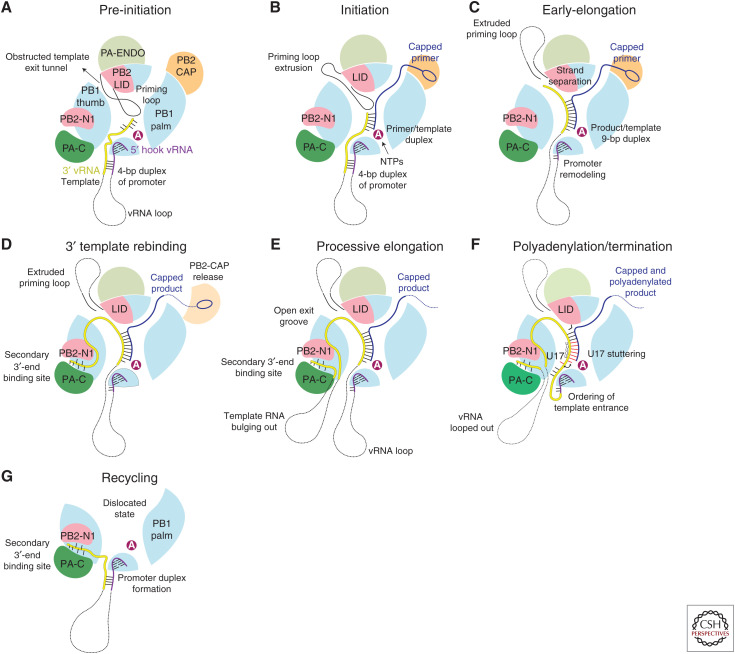
References
-
- Byrn RA, Jones SM, Bennett HB, Bral C, Clark MP, Jacobs MD, Kwong AD, Ledeboer MW, Leeman JR, McNeil CF, et al. 2015. Preclinical activity of VX-787, a first-in-class, orally bioavailable inhibitor of the influenza virus polymerase PB2 subunit. Antimicrob Agents Chemother 59: 1569–1582. 10.1128/AAC.04623-14 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous