

In Vitro Impact of FSH Glycosylation Variants on FSH Receptor-stimulated Signal Transduction and Functional Selectivity
- PMID: 32342021
- PMCID: PMC7175721
- DOI: 10.1210/jendso/bvaa019
In Vitro Impact of FSH Glycosylation Variants on FSH Receptor-stimulated Signal Transduction and Functional Selectivity
Abstract
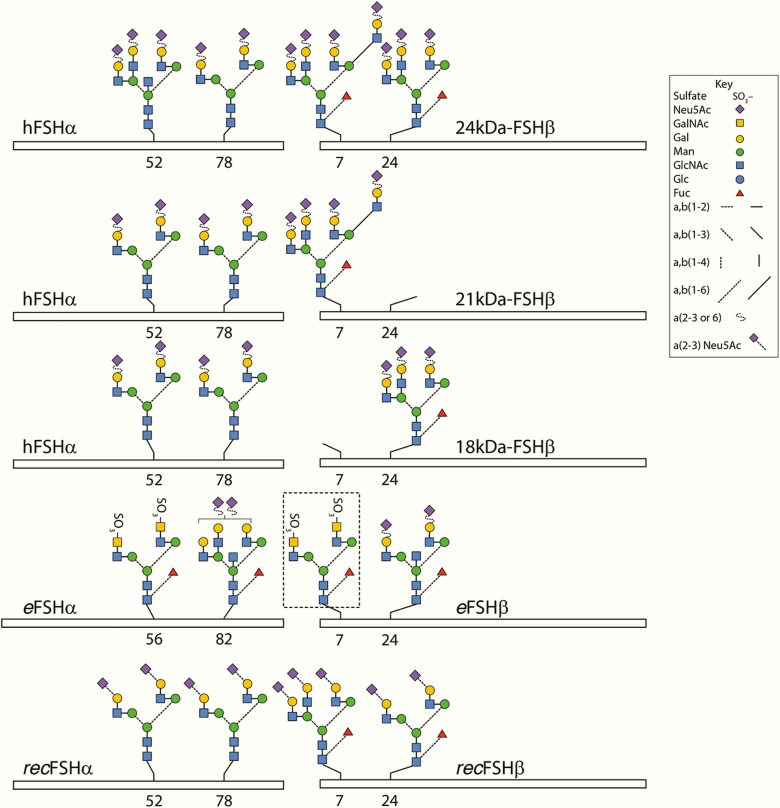
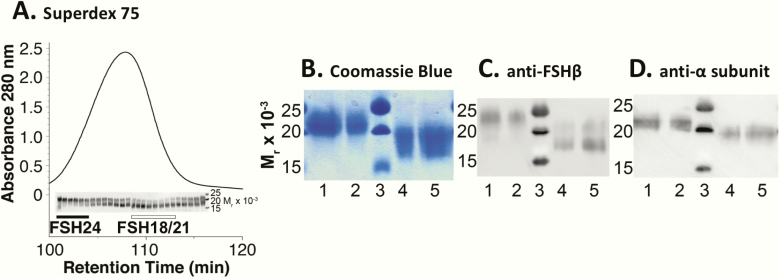
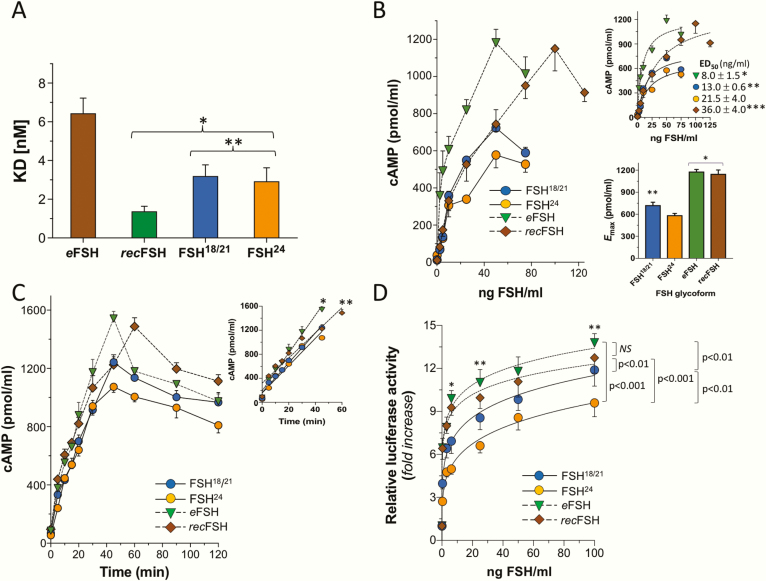
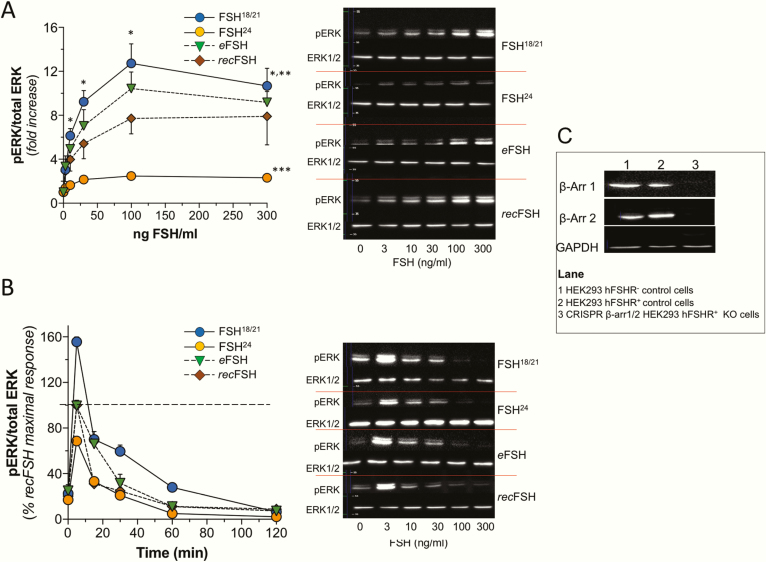
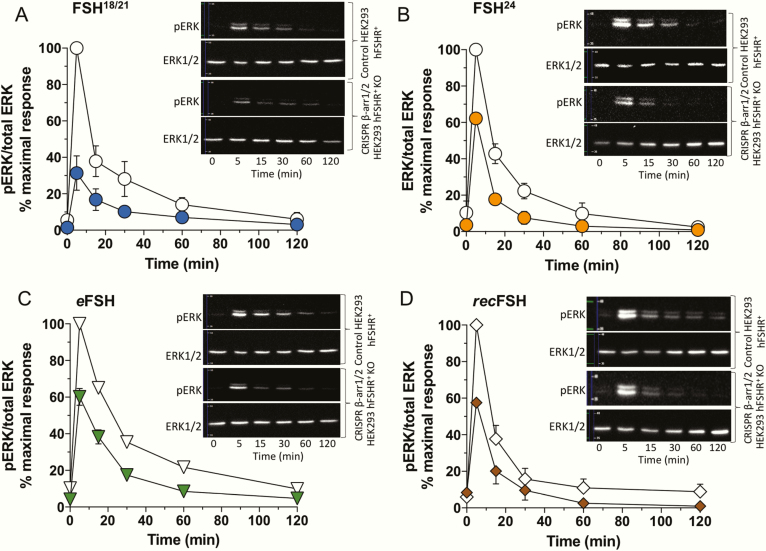
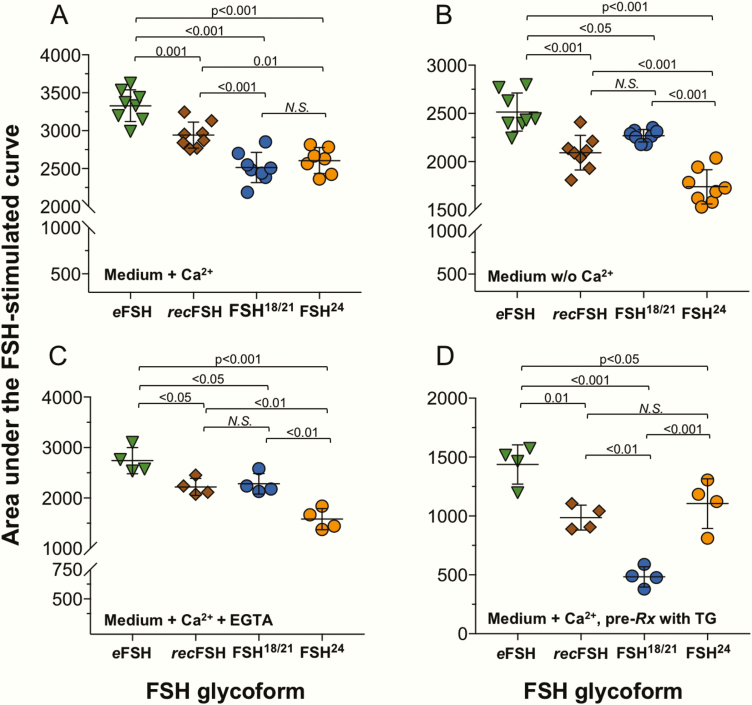
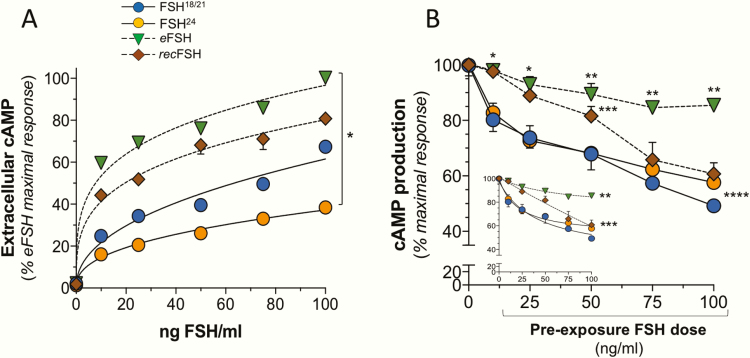
FSH exists as different glycoforms that differ in glycosylation of the hormone-specific β-subunit. Tetra-glycosylated FSH (FSH24) and hypo-glycosylated FSH (FSH18/21) are the most abundant glycoforms found in humans. Employing distinct readouts in HEK293 cells expressing the FSH receptor, we compared signaling triggered by human pituitary FSH preparations (FSH18/21 and FSH24) as well as by equine FSH (eFSH), and human recombinant FSH (recFSH), each exhibiting distinct glycosylation patterns. The potency in eliciting cAMP production was greater for eFSH than for FSH18/21, FSH24, and recFSH, whereas in the ERK1/2 activation readout, potency was highest for FSH18/21 followed by eFSH, recFSH, and FSH24. In β-arrestin1/2 CRISPR/Cas9 HEK293-KO cells, FSH18/21 exhibited a preference toward β-arrestin-mediated ERK1/2 activation as revealed by a drastic decrease in pERK during the first 15-minute exposure to this glycoform. Exposure of β-arrestin1/2 KO cells to H89 additionally decreased pERK1/2, albeit to a significantly lower extent in response to FSH18/21. Concurrent silencing of β-arrestin and PKA signaling, incompletely suppressed pERK response to FSH glycoforms, suggesting that pathways other than those dependent on Gs-protein and β-arrestins also contribute to FSH-stimulated pERK1/2. All FSH glycoforms stimulated intracellular Ca2+ (iCa2+) accumulation through both influx from Ca2+ channels and release from intracellular stores; however, iCa2+ in response to FSH18/21 depended more on the latter, suggesting differences in mechanisms through which glycoforms promote iCa2+ accumulation. These data indicate that FSH glycosylation plays an important role in defining not only the intensity but also the functional selectivity for the mechanisms leading to activation of distinct signaling cascades.
Keywords: biased agonism; follicle-stimulating hormone; follicle-stimulating hormone receptor; functional selectivity; glycosylation; macroheterogeneity; signal transduction.
© Endocrine Society 2020.
Figures

Similar articles
-
Hypo-glycosylated hFSH drives ovarian follicular development more efficiently than fully-glycosylated hFSH: enhanced transcription and PI3K and MAPK signaling.Hum Reprod. 2021 Jun 18;36(7):1891-1906. doi: 10.1093/humrep/deab135. Hum Reprod. 2021. PMID: 34059912 Free PMC article.
-
Differential effects of follicle-stimulating hormone glycoforms on the transcriptome profile of cultured rat granulosa cells as disclosed by RNA-seq.PLoS One. 2024 Jun 6;19(6):e0293688. doi: 10.1371/journal.pone.0293688. eCollection 2024. PLoS One. 2024. PMID: 38843139 Free PMC article.
-
Differential effects of follicle-stimulating hormone glycoforms on the transcriptome profile of cultured rat granulosa cells as disclosed by RNA-seq.bioRxiv [Preprint]. 2023 Oct 20:2023.10.18.562995. doi: 10.1101/2023.10.18.562995. bioRxiv. 2023. Update in: PLoS One. 2024 Jun 6;19(6):e0293688. doi: 10.1371/journal.pone.0293688. PMID: 37905087 Free PMC article. Updated. Preprint.
-
Biased signaling by human follicle-stimulating hormone variants.Pharmacol Ther. 2025 Apr;268:108821. doi: 10.1016/j.pharmthera.2025.108821. Epub 2025 Feb 15. Pharmacol Ther. 2025. PMID: 39961417 Review.
-
Structural features of mammalian gonadotropins.Mol Cell Endocrinol. 1996 Dec 20;125(1-2):3-19. doi: 10.1016/s0303-7207(96)03945-7. Mol Cell Endocrinol. 1996. PMID: 9027339 Review.
Cited by
-
Hypo-glycosylated hFSH drives ovarian follicular development more efficiently than fully-glycosylated hFSH: enhanced transcription and PI3K and MAPK signaling.Hum Reprod. 2021 Jun 18;36(7):1891-1906. doi: 10.1093/humrep/deab135. Hum Reprod. 2021. PMID: 34059912 Free PMC article.
-
Differential effects of follicle-stimulating hormone glycoforms on the transcriptome profile of cultured rat granulosa cells as disclosed by RNA-seq.PLoS One. 2024 Jun 6;19(6):e0293688. doi: 10.1371/journal.pone.0293688. eCollection 2024. PLoS One. 2024. PMID: 38843139 Free PMC article.
-
New Human Follitropin Preparations: How Glycan Structural Differences May Affect Biochemical and Biological Function and Clinical Effect.Front Endocrinol (Lausanne). 2021 Mar 19;12:636038. doi: 10.3389/fendo.2021.636038. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 33815292 Free PMC article. Review.
-
Differential effects of follicle-stimulating hormone glycoforms on the transcriptome profile of cultured rat granulosa cells as disclosed by RNA-seq.bioRxiv [Preprint]. 2023 Oct 20:2023.10.18.562995. doi: 10.1101/2023.10.18.562995. bioRxiv. 2023. Update in: PLoS One. 2024 Jun 6;19(6):e0293688. doi: 10.1371/journal.pone.0293688. PMID: 37905087 Free PMC article. Updated. Preprint.
-
Biased signaling in naturally occurring mutations of G protein-coupled receptors associated with diverse human diseases.Biochim Biophys Acta Mol Basis Dis. 2021 Jan 1;1867(1):165973. doi: 10.1016/j.bbadis.2020.165973. Epub 2020 Sep 17. Biochim Biophys Acta Mol Basis Dis. 2021. PMID: 32949766 Free PMC article. Review.
References
-
- Baenziger JU. Glycoprotein hormone GalNAc-4-sulphotransferase. Biochem Soc Trans. 2003;31(2):326–330. - PubMed
-
- Chappel SC, Ulloa-Aguirre A, Coutifaris C. Biosynthesis and secretion of follicle-stimulating hormone. Endocr Rev. 1983;4(2):179–211. - PubMed
-
- Pierce JG, Parsons TF. Glycoprotein hormones: structure and function. Annu Rev Biochem. 1981;50:465–495. - PubMed
-
- Bousfield GR, Butnev VY, Gotschall RR, Baker VL, Moore WT. Structural features of mammalian gonadotropins. Mol Cell Endocrinol. 1996;125(1-2):3–19. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous