

nNOS-CAPON blockers produce anxiolytic effects by promoting synaptogenesis in chronic stress-induced animal models of anxiety
- PMID: 32343840
- PMCID: PMC7393190
- DOI: 10.1111/bph.15084
nNOS-CAPON blockers produce anxiolytic effects by promoting synaptogenesis in chronic stress-induced animal models of anxiety
Abstract
Background and purpose: Anxiety disorder is a common mental health disorder. However, there are few safe and fast-acting anxiolytic drugs available that can treat anxiety disorder. We previously demonstrated that the interaction of neuronal NOS (nNOS) with its carboxy-terminal PDZ ligand (CAPON) is involved in regulating anxiety-related behaviours. Here, we further investigated the anxiolytic effects of nNOS-CAPON disruptors in chronic stress-induced anxiety in animals.
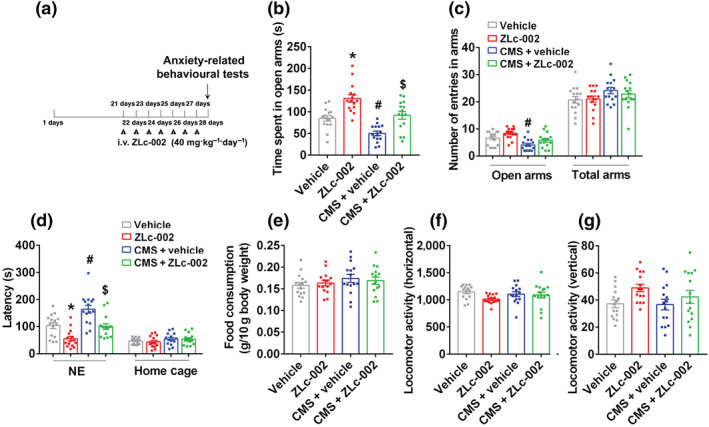
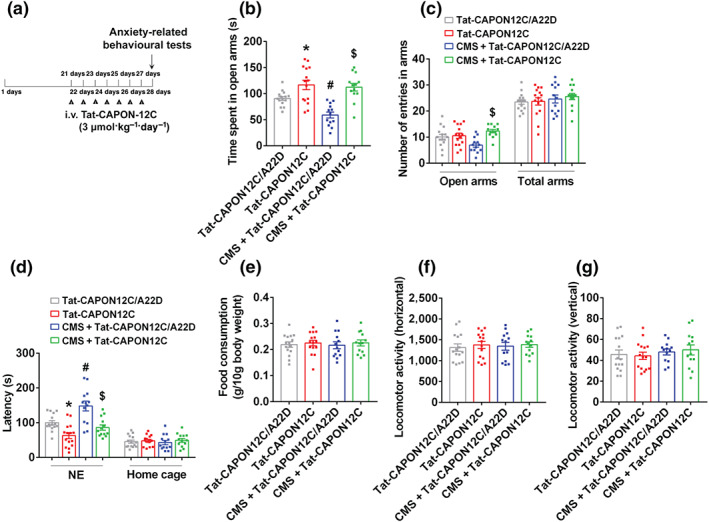
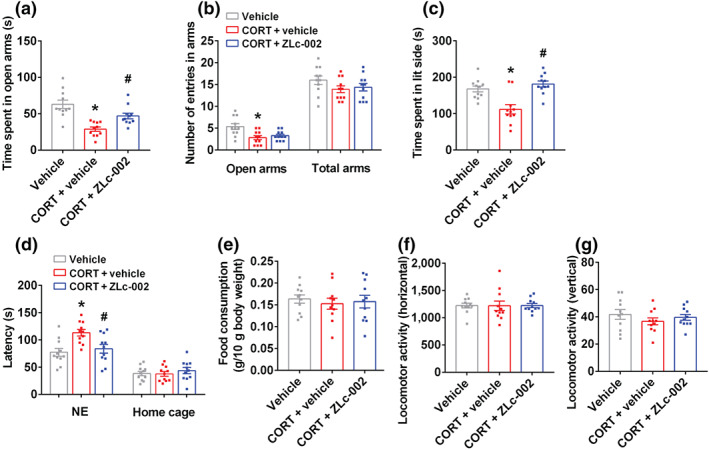
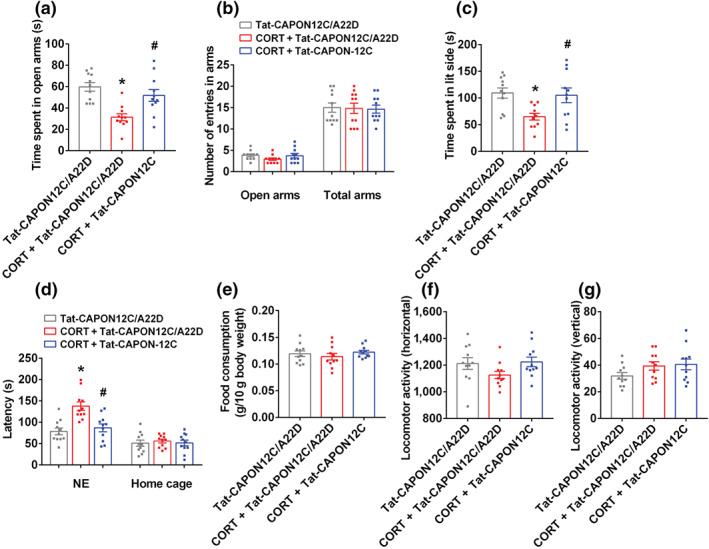
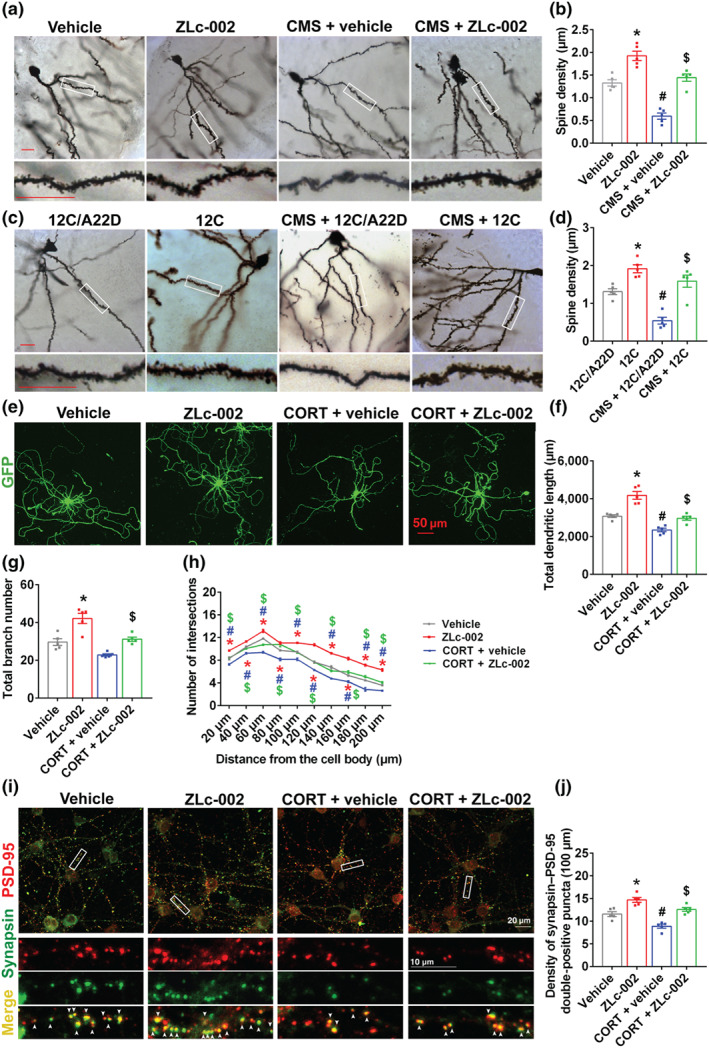
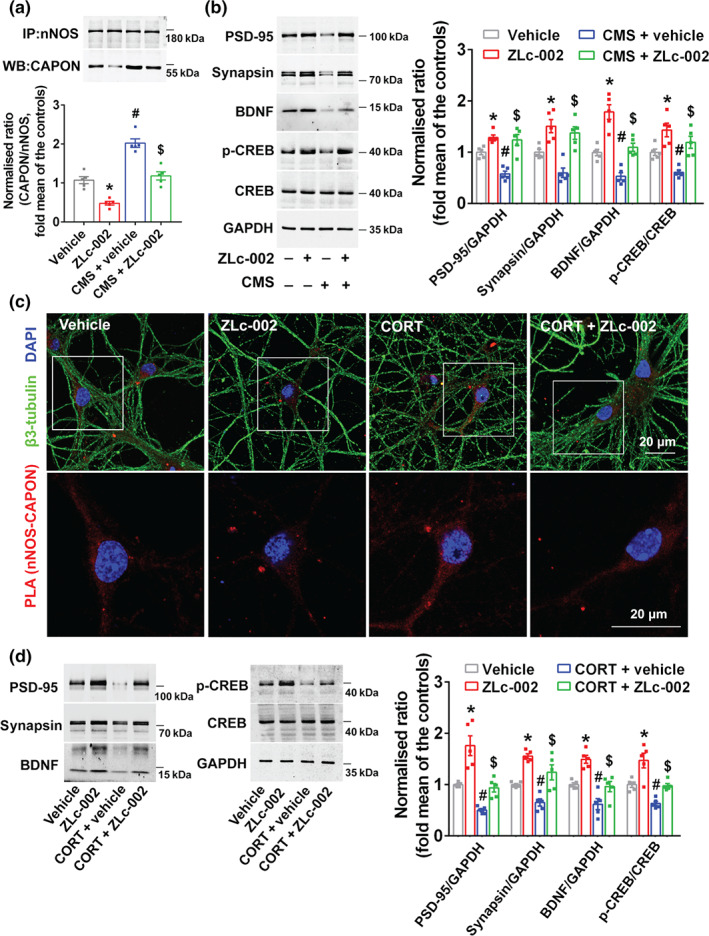
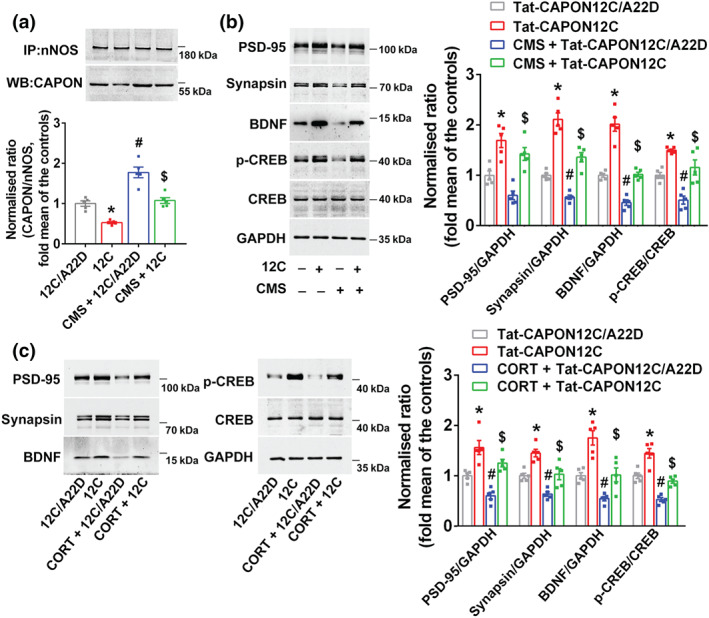
Experimental approach: Mice were intravenously treated with nNOS-CAPON disruptors, ZLc-002 or Tat-CAPON12C, at the last week of chronic mild stress (CMS) exposure. We also infused corticosterone (CORT) into the hippocampus of mice to model anxiety behaviours and also delivered ZLc-002 or Tat-CAPON12C on the last week of chronic CORT treatment via pre-implanted cannula. Anxiety-related behaviours were examined using elevated plus maze, open field, novelty-suppressed feeding and light-dark (LD) tests. The level of nNOS-CAPON interaction was determined by co-immunoprecipitation (CO-IP) and proximity ligation assay (PLA). The neural mechanisms underlying the behavioural effects of nNOS-CAPON uncoupling in anxiety animal models were assessed by western blot, immunofluorescence and Golgi-Cox staining.
Key results: ZLc-002 and Tat-CAPON12C reversed CMS- or CORT-induced anxiety-related behaviours. ZLc-002 and Tat-CAPON12C increased synaptogenesis along with improved dendritic remodelling in CMS mice or CORT-treated cultured neurons. Meanwhile, blocking nNOS-CAPON interaction significantly activated the cAMP response element-binding protein (CREB)-brain-derived neurotrophic factor (BDNF) pathway, which is associated with synaptic plasticity.
Conclusion and implications: Collectively, these results provide evidence for the anxiolytic effects of nNOS-CAPON uncouplers and their underlying mechanisms in anxiety disorders.
© 2020 The British Pharmacological Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
-
- Alexander, S. P. H. , Roberts, R. E. , Broughton, B. R. S. , Sobey, C. G. , George, C. H. , Stanford, S. C. , … Ahluwalia, A. (2018). Goals and practicalities of immunoblotting and immunohistochemistry: A guide for submission to the British Journal of Pharmacology . British Journal of Pharmacology, 175(3), 407–411. 10.1111/bph.14112 - DOI - PMC - PubMed
-
- Almandoz‐Gil, L. , Persson, E. , Lindström, V. , Ingelsson, M. , Erlandsson, A. , & Bergström, J. (2018). In situ proximity ligation assay reveals co‐localization of alpha‐synuclein and SNARE proteins in murine primary neurons. Frontiers in Neurology, 9, 180 10.3389/fneur.2018.00180 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 2016YFA0501001/National Key R&D Program of China
- 2016YFC1306703/National Key R&D Program of China
- 81401121/National Natural Science Foundation of China
- 31530091/National Natural Science Foundation of China
- 31671107/National Natural Science Foundation of China
- 31970951/National Natural Science Foundation of China
- BK20140366/Natural Science Foundation of Jiangsu Province
- BK20170021/Natural Science Foundation of Jiangsu Province
- Fundamental Research Funds for the Central Universities
- 2016QNRC001/Young Elite Scientists Sponsorship Program by CAST
- 'Zhong Ying Young Scholar' project of Cyrus Tang Foundation
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
