

Pin1 promotes pancreatic cancer progression and metastasis by activation of NF-κB-IL-18 feedback loop
- PMID: 32347623
- PMCID: PMC7260075
- DOI: 10.1111/cpr.12816
Pin1 promotes pancreatic cancer progression and metastasis by activation of NF-κB-IL-18 feedback loop
Abstract
Objectives: Accumulated evidence suggests that Pin1 contributes to oncogenesis of diverse cancers. However, the underlying mechanism of oncogenic function of Pin1 in PDAC requires further exploration.
Materials and methods: IHC was performed using PDAC tissues. Western blot, PCR, immunofluorescence and transwell were performed using cell lines. GSEA were applied for possible downstream pathways. ChIP assay and dual luciferase were used for assessment of transcriptional activity.
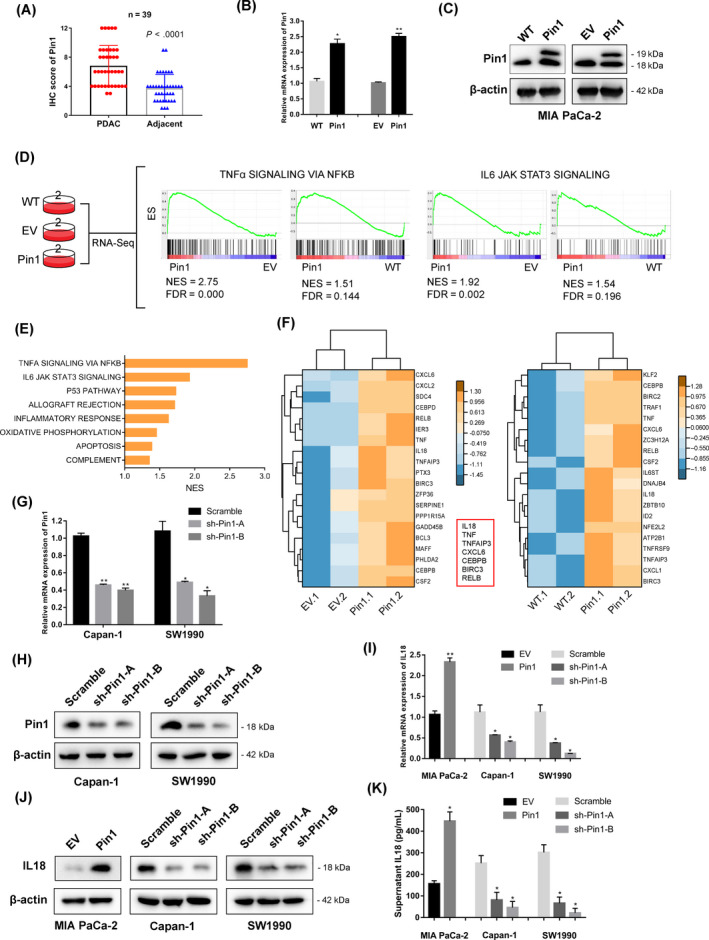
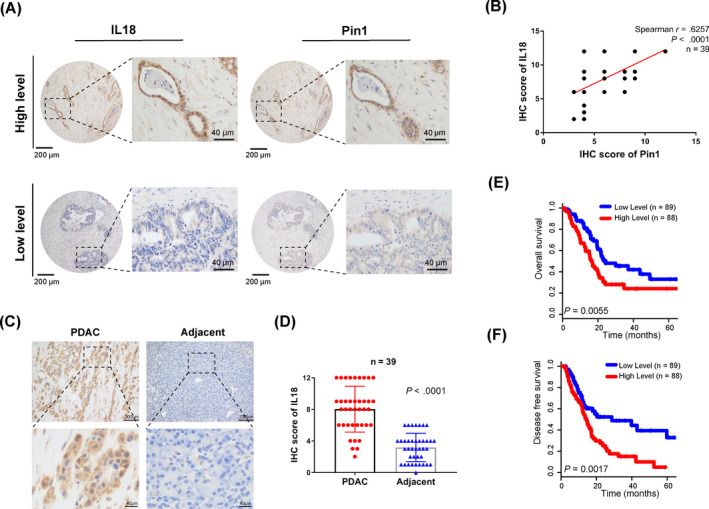
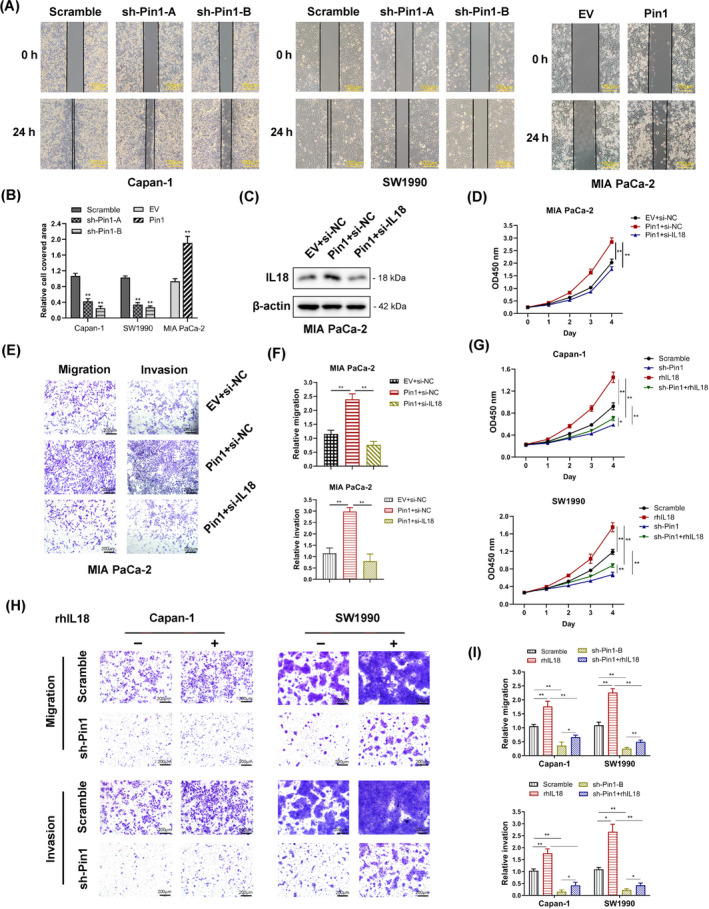
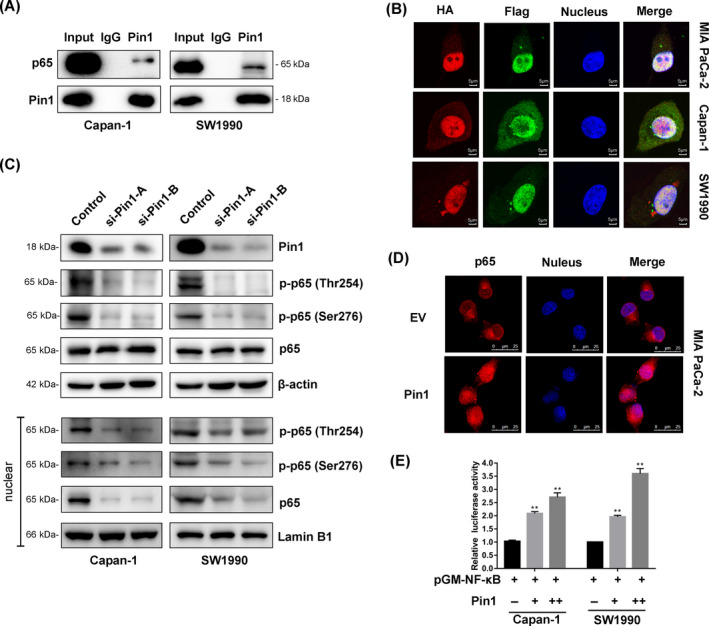
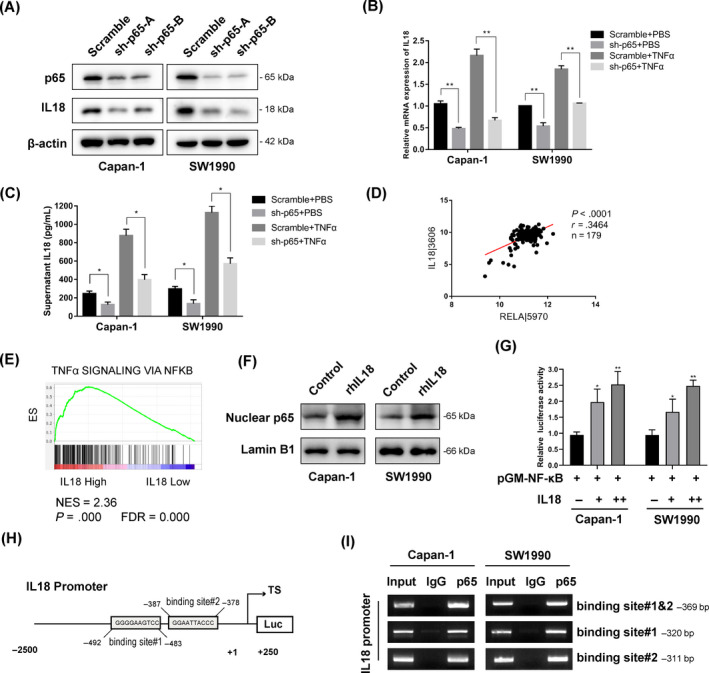
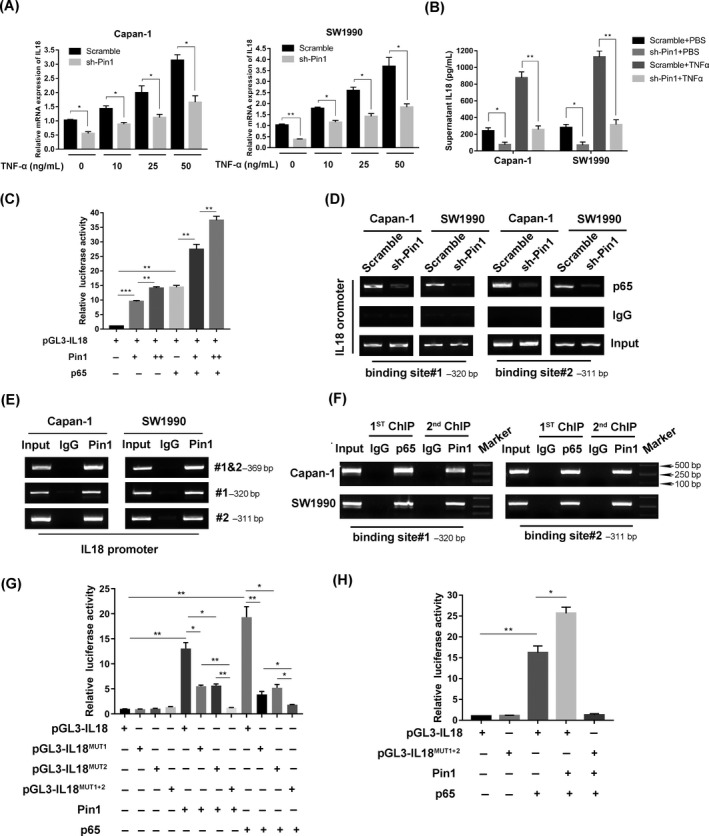
Results: Both Pin1 and IL-18 levels are increased in primary PDAC tissues and that their levels are positively correlated. High expression of IL-18 is a predictor of poor prognoses. Pin1 promoted pancreatic cancer cell proliferation and motility by increasing IL-18 expression, while Pin1 knockdown also inhibited the tumour-promoting effect of IL-18. Both Pin1 and IL-18 could enhance the NFκB activity in pancreatic cancer cells. When bound to the p65 protein, Pin1 promoted p65 phosphorylation and its nuclear translocation. In the nucleus, Pin1 and p65 simultaneously bound to the IL-18 promoter and enhanced IL-18 transcription. In addition, recruitment of p65 to the IL-18 promoter was decreased in Pin1-silenced cells.
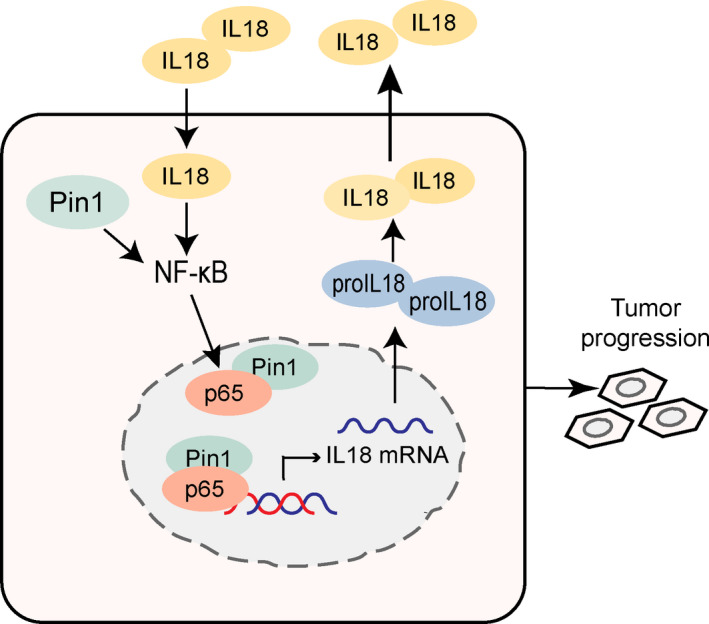
Conclusions: Our study improves the understanding of Pin1 in tumour-promoting inflammation in PDAC, which is a hallmark of cancer; Pin1 interacted with p65 in PDAC and enhanced NF-κB signalling and downstream transcriptional activation of IL-18, with increased IL-18 continuously activating NF-κB signalling, which then forms a positive feedback loop.
Keywords: PIN1; gene expression; interleukin 18; pancreatic cancer.
© 2020 The Authors. Cell Proliferation published by John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Siegel RL, Miller KD, Jemal A. Cancer statistics, 2018. CA Cancer J Clin. 2018;68(1):7‐30. - PubMed
-
- Midha S, Chawla S, Garg PK. Modifiable and non‐modifiable risk factors for pancreatic cancer: a review. Cancer Lett. 2016;381(1):269‐277. - PubMed
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646‐674. - PubMed
-
- Greten FR, Eckmann L, Greten TF et al IKKbeta links inflammation and tumorigenesis in a mouse model of colitis‐associated cancer. Cell. 2004;118(3):285‐296. - PubMed
-
- Ghosh S, Karin M. Missing pieces in the NF‐kappaB puzzle. Cell. 2002;109(Suppl):S81‐96. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
